

Microtubule-based perception of mechanical conflicts controls plant organ morphogenesis
- PMID: 35138892
- PMCID: PMC8827652
- DOI: 10.1126/sciadv.abm4974
Microtubule-based perception of mechanical conflicts controls plant organ morphogenesis
Abstract
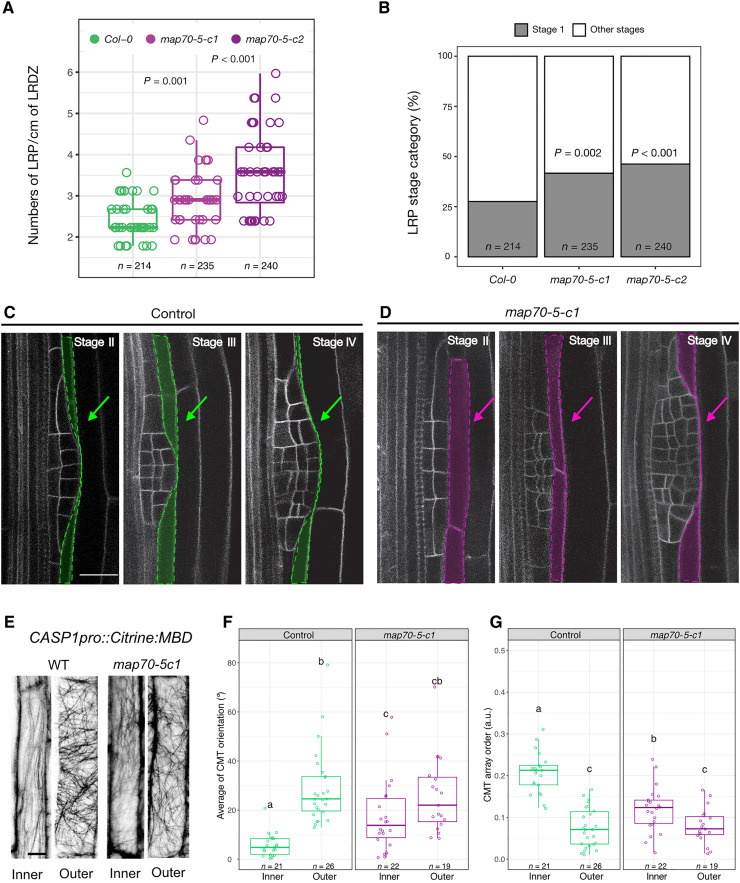
Precise coordination between cells and tissues is essential for differential growth in plants. During lateral root formation in Arabidopsis thaliana, the endodermis is actively remodeled to allow outgrowth of the new organ. Here, we show that microtubule arrays facing lateral root founder cells display a higher order compared to arrays on the opposite side of the same cell, and this asymmetry is required for endodermal remodeling and lateral root initiation. We identify that MICROTUBULE ASSOCIATED PROTEIN 70-5 (MAP70-5) is necessary for the establishment of this spatially defined microtubule organization and endodermis remodeling and thus contributes to lateral root morphogenesis. We propose that MAP70-5 and cortical microtubule arrays in the endodermis integrate the mechanical signals generated by lateral root outgrowth, facilitating the channeling of organogenesis.
Figures




Similar articles
-
Cytoskeleton Dynamics Are Necessary for Early Events of Lateral Root Initiation in Arabidopsis.Curr Biol. 2019 Aug 5;29(15):2443-2454.e5. doi: 10.1016/j.cub.2019.06.039. Epub 2019 Jul 18. Curr Biol. 2019. PMID: 31327713
-
AtMYB93 is an endodermis-specific transcriptional regulator of lateral root development in arabidopsis.Plant Signal Behav. 2014;9(10):e970406. doi: 10.4161/15592316.2014.970406. Plant Signal Behav. 2014. PMID: 25482809 Free PMC article.
-
Spatiotemporal Pattern of Ectopic Cell Divisions Contribute to Mis-Shaped Phenotype of Primary and Lateral Roots of katanin1 Mutant.Front Plant Sci. 2020 Jun 9;11:734. doi: 10.3389/fpls.2020.00734. eCollection 2020. Front Plant Sci. 2020. PMID: 32582258 Free PMC article.
-
Lateral root initiation in Arabidopsis thaliana: a force awakens.F1000Prime Rep. 2015 Mar 3;7:32. doi: 10.12703/P7-32. eCollection 2015. F1000Prime Rep. 2015. PMID: 25926983 Free PMC article. Review.
-
Microtubule organization and microtubule-associated proteins in plant cells.Int Rev Cell Mol Biol. 2014;312:1-52. doi: 10.1016/B978-0-12-800178-3.00001-4. Int Rev Cell Mol Biol. 2014. PMID: 25262237 Review.
Cited by
-
Arabidopsis floral buds are locked through stress-induced sepal tip curving.Nat Plants. 2024 Aug;10(8):1258-1266. doi: 10.1038/s41477-024-01760-6. Epub 2024 Jul 26. Nat Plants. 2024. PMID: 39060423
-
Open quantum systems theory of ultraweak ultraviolet photon emissions: Revisiting Gurwitsch's onion experiment as a prototype for quantum biology.Comput Struct Biotechnol J. 2024 Nov 29;26:78-91. doi: 10.1016/j.csbj.2024.11.030. eCollection 2024 Dec. Comput Struct Biotechnol J. 2024. PMID: 39717158 Free PMC article. Review.
-
Microtubule Regulation in Plants: From Morphological Development to Stress Adaptation.Biomolecules. 2023 Mar 30;13(4):627. doi: 10.3390/biom13040627. Biomolecules. 2023. PMID: 37189374 Free PMC article. Review.
-
HY5 inhibits lateral root initiation in Arabidopsis through negative regulation of the microtubule-stabilizing protein TPXL5.Plant Cell. 2023 Mar 15;35(3):1092-1109. doi: 10.1093/plcell/koac358. Plant Cell. 2023. PMID: 36512471 Free PMC article.
-
Microtubule-associated protein SlMAP70 interacts with IQ67-domain protein SlIQD21a to regulate fruit shape in tomato.Plant Cell. 2023 Nov 30;35(12):4266-4283. doi: 10.1093/plcell/koad231. Plant Cell. 2023. PMID: 37668409 Free PMC article.
References
-
- Hamant O., Widespread mechanosensing controls the structure behind the architecture in plants. Curr. Opin. Plant Biol. 16, 654–660 (2013). - PubMed
-
- Hamant O., Heisler M. G., Jönsson H., Krupinski P., Uyttewaal M., Bokov P., Corson F., Sahlin P., Boudaoud A., Meyerowitz E. M., Couder Y., Traas J., Developmental patterning by mechanical signals in Arabidopsis. Science 322, 1650–1655 (2008). - PubMed
-
- Hamant O., Saunders T. E., Shaping organs: Shared structural principles across kingdoms. Annu. Rev. Cell Dev. Biol. 36, 385–410 (2020). - PubMed
LinkOut - more resources
Full Text Sources
