

Innate Sensors Trigger Regulated Cell Death to Combat Intracellular Infection
- PMID: 35138947
- PMCID: PMC9614550
- DOI: 10.1146/annurev-immunol-101320-011235
Innate Sensors Trigger Regulated Cell Death to Combat Intracellular Infection
Abstract
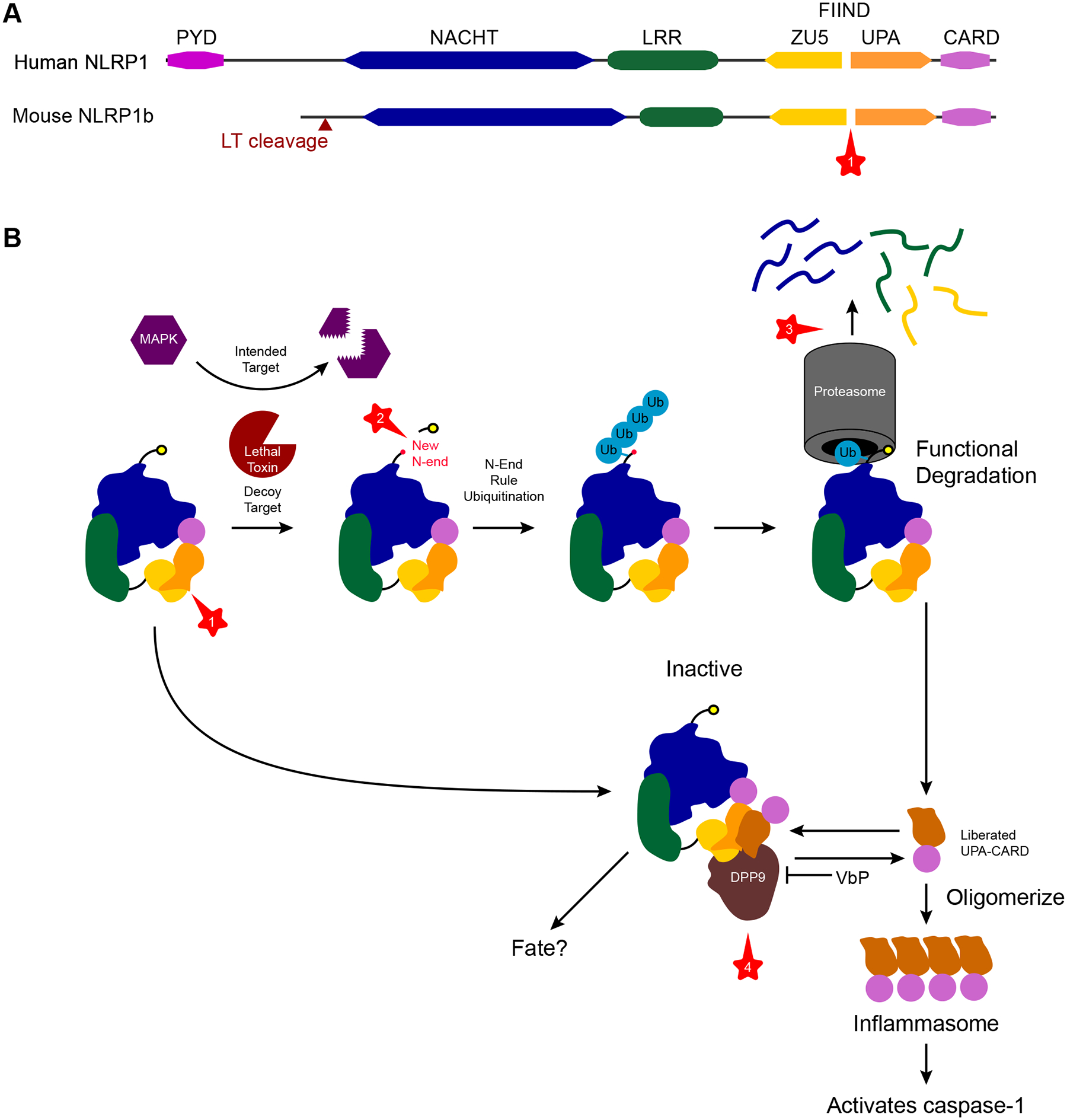
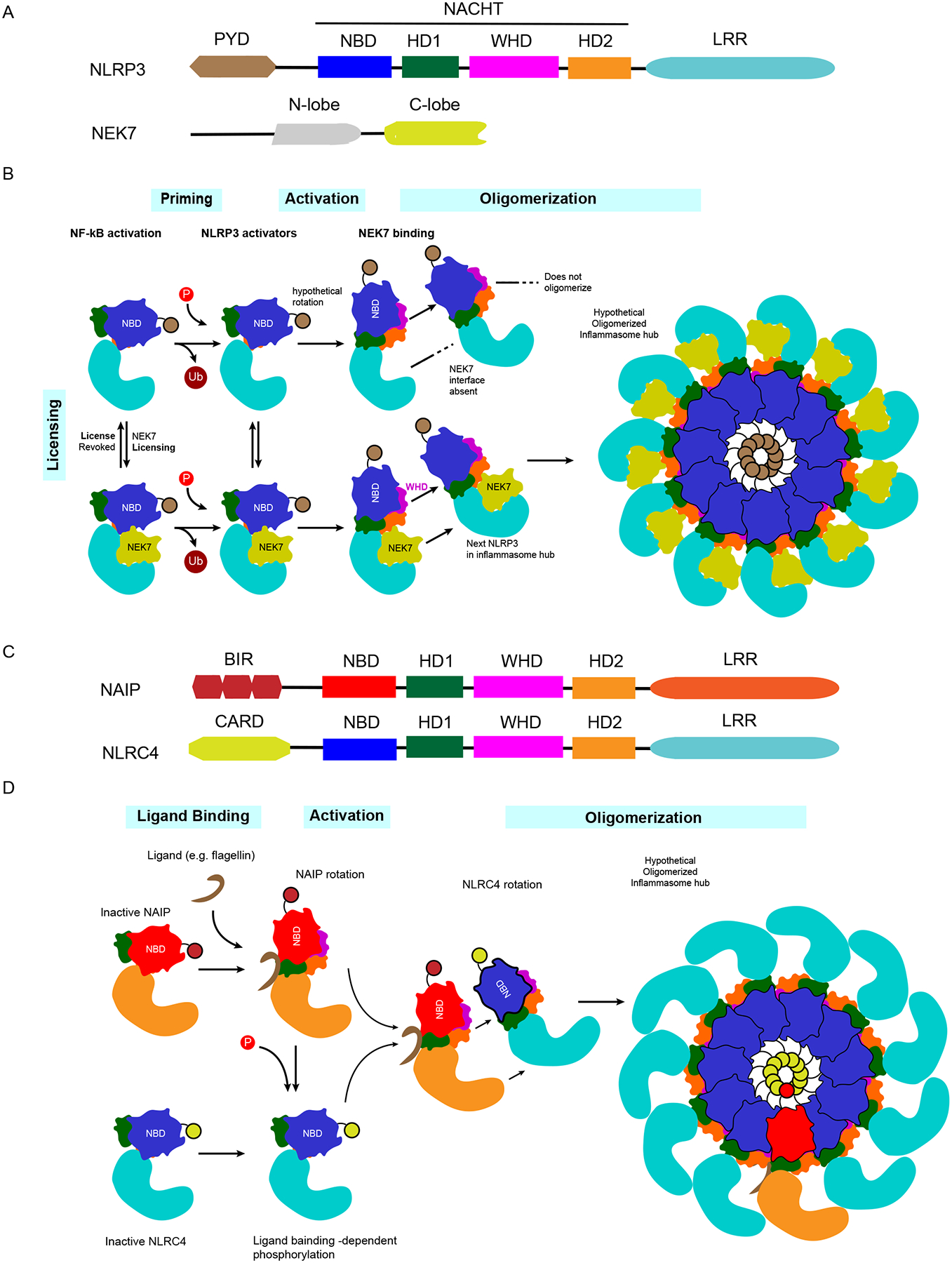
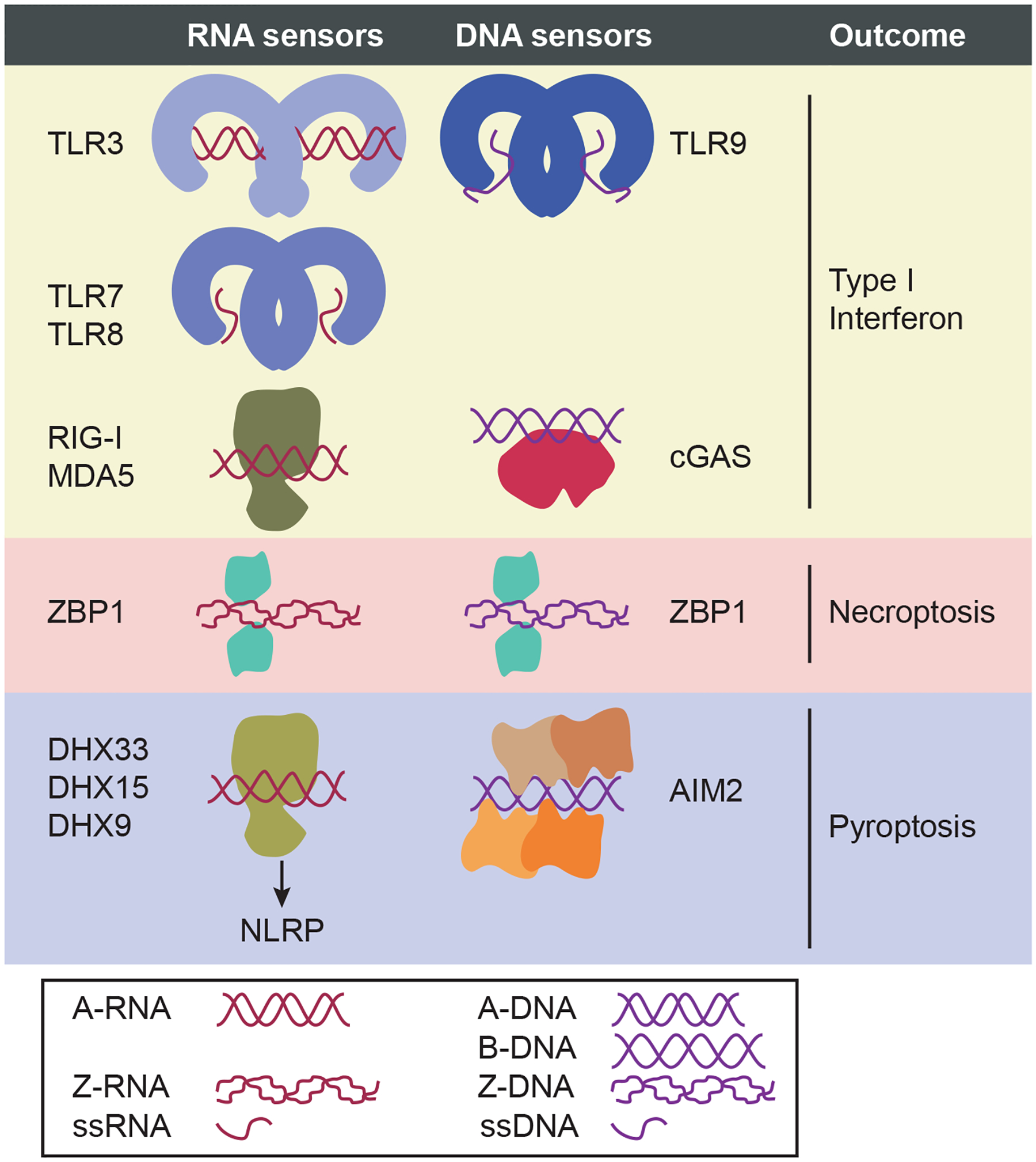
Intracellular pathogens pose a significant threat to animals. In defense, innate immune sensors attempt to detect these pathogens using pattern recognition receptors that either directly detect microbial molecules or indirectly detect their pathogenic activity. These sensors trigger different forms of regulated cell death, including pyroptosis, apoptosis, and necroptosis, which eliminate the infected host cell niche while simultaneously promoting beneficial immune responses. These defenses force intracellular pathogens to evolve strategies to minimize or completely evade the sensors. In this review, we discuss recent advances in our understanding of the cytosolic pattern recognition receptors that drive cell death, including NLRP1, NLRP3, NLRP6, NLRP9, NLRC4, AIM2, IFI16, and ZBP1.
Keywords: caspase; inflammasome; innate immunity; necroptosis; pattern recognition receptors; pyroptosis.
Figures
References
-
- Van Valen L 1973. A new evolutionary law. Evol. Theory 1:1–30
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
