

Infection with the enteric pathogen C. rodentium promotes islet-specific autoimmunity by activating a lymphatic route from the gut to pancreatic lymph node
- PMID: 35140345
- PMCID: PMC9038524
- DOI: 10.1038/s41385-022-00490-2
Infection with the enteric pathogen C. rodentium promotes islet-specific autoimmunity by activating a lymphatic route from the gut to pancreatic lymph node
Abstract
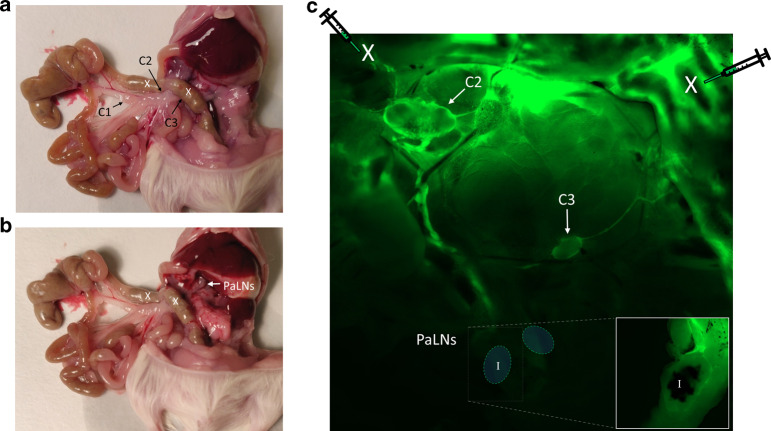
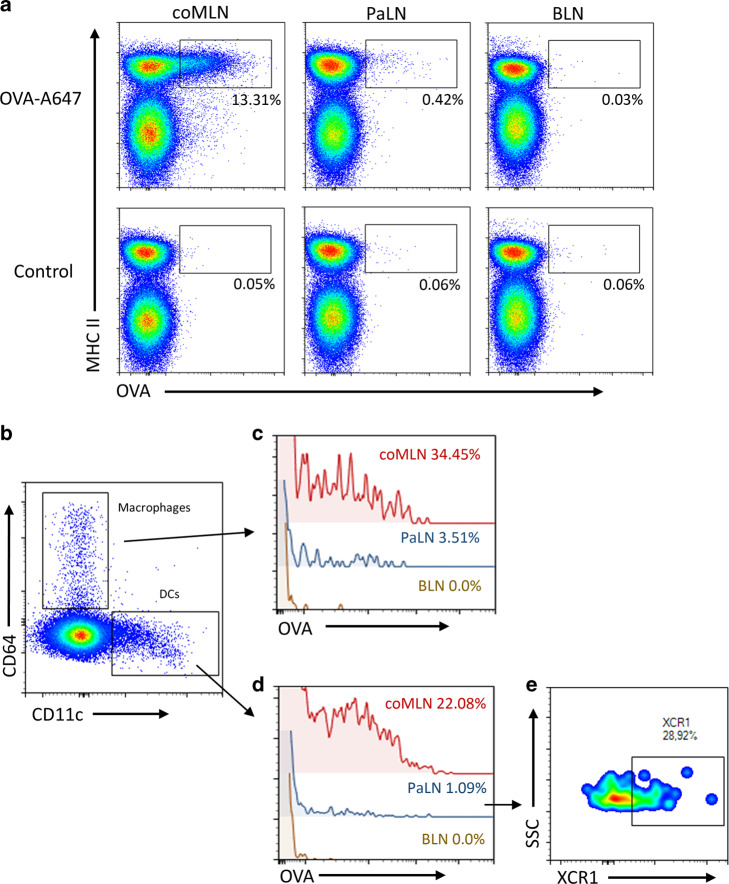
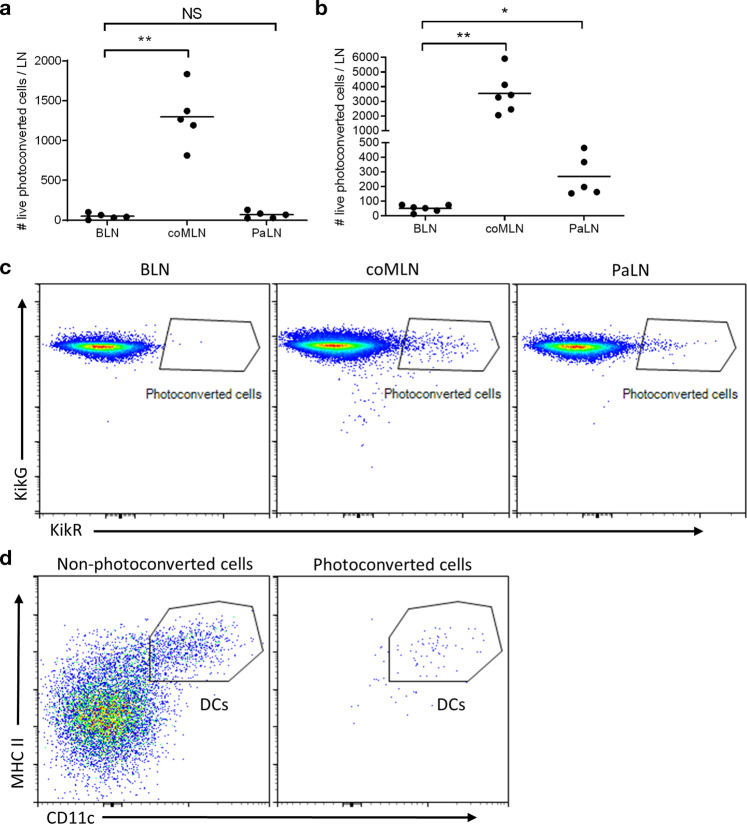
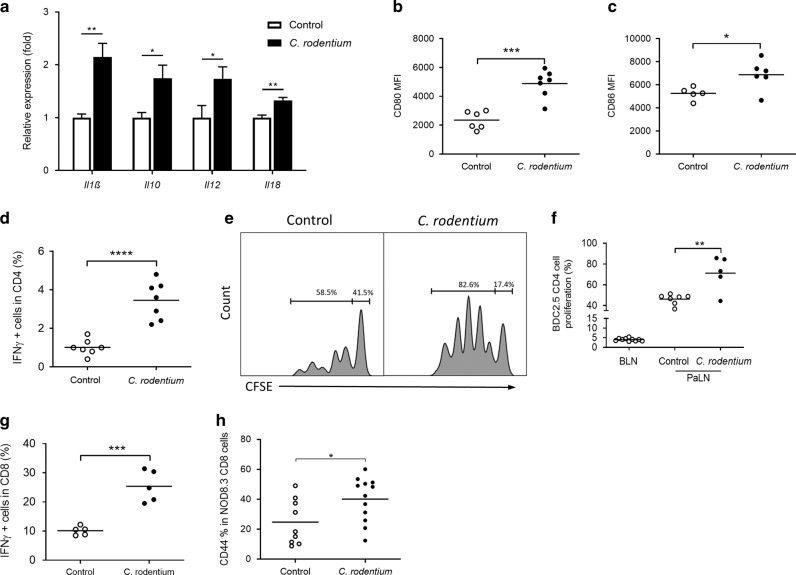
In nonobese diabetic (NOD) mice, C. rodentium promotes priming of islet-specific T-cells in pancreatic lymph nodes (PaLN), which is a critical step in initiation and perpetuation of islet-autoimmunity. To investigate mechanisms by which C. rodentium promotes T-cell priming in PaLN, we used fluorescent imaging of lymphatic vasculature emanating from colon, followed dendritic cell (DC) migration from colon using photoconvertible-reporter mice, and evaluated the translocation of bacteria to lymph nodes with GFP-C. rodentium and in situ hybridization of bacterial DNA. Fluorescent dextran injected in the colon wall accumulated under subcapsular sinus of PaLN indicating the existence of a lymphatic route from colon to PaLN. Infection with C. rodentium induced DC migration from colon to PaLN and bacterial DNA was detected in medullary sinus and inner cortex of PaLN. Following infection with GFP-C. rodentium, fluorescence appeared in macrophages and gut-derived (CD103+) and resident (CD103-/XCR1+) DC, indicating transportation of bacteria from colon to PaLN both by DC and by lymph itself. This induced proinflammatory cytokine transcripts, activation of DC and islet-specific T-cells in PaLN of NOD mice. Our findings demonstrate the existence of a direct, enteric pathogen-activated route for lymph, cells, and bacteria from colon, which promotes activation of islet-specific T-cells in PaLN.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
