

Systematic Phenotyping and Characterization of the 3xTg-AD Mouse Model of Alzheimer's Disease
- PMID: 35140584
- PMCID: PMC8818877
- DOI: 10.3389/fnins.2021.785276
Systematic Phenotyping and Characterization of the 3xTg-AD Mouse Model of Alzheimer's Disease
Abstract
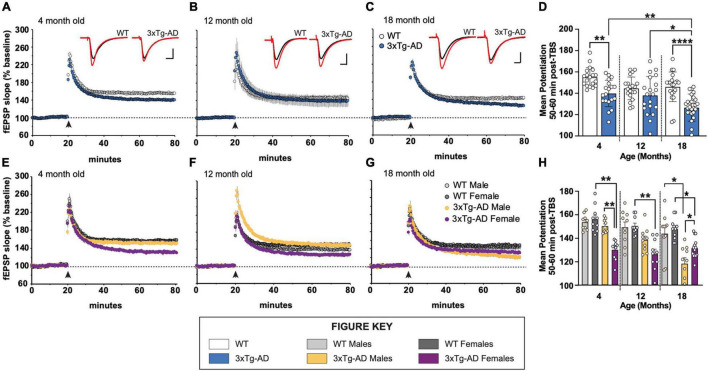
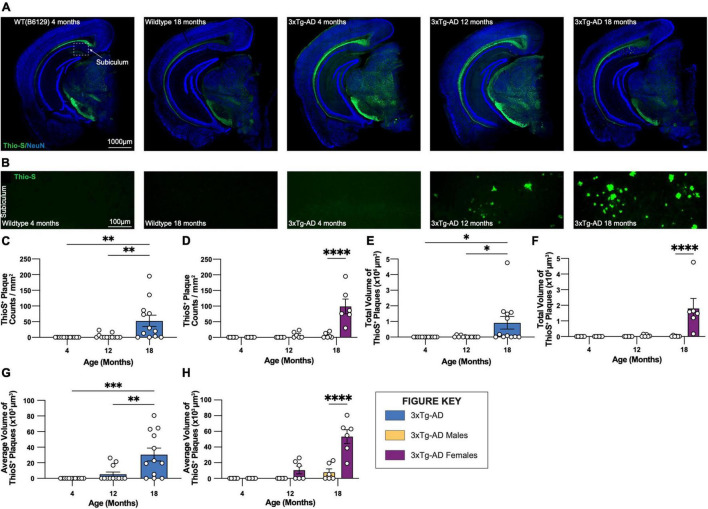
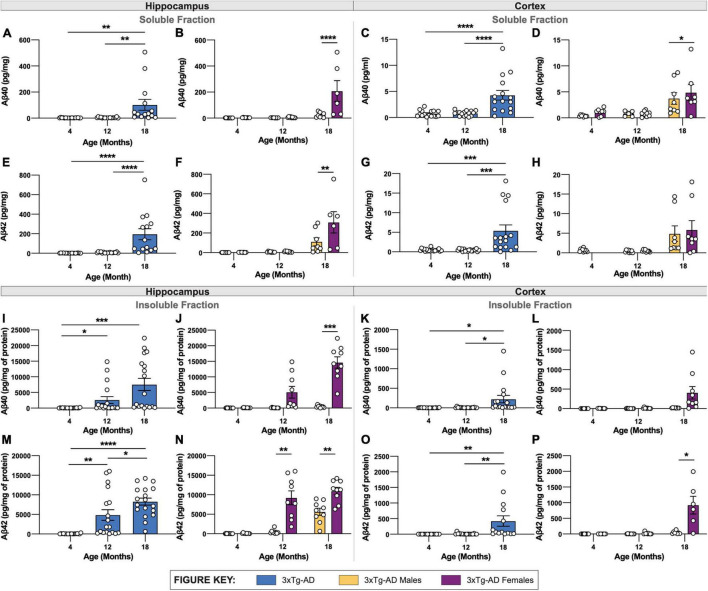
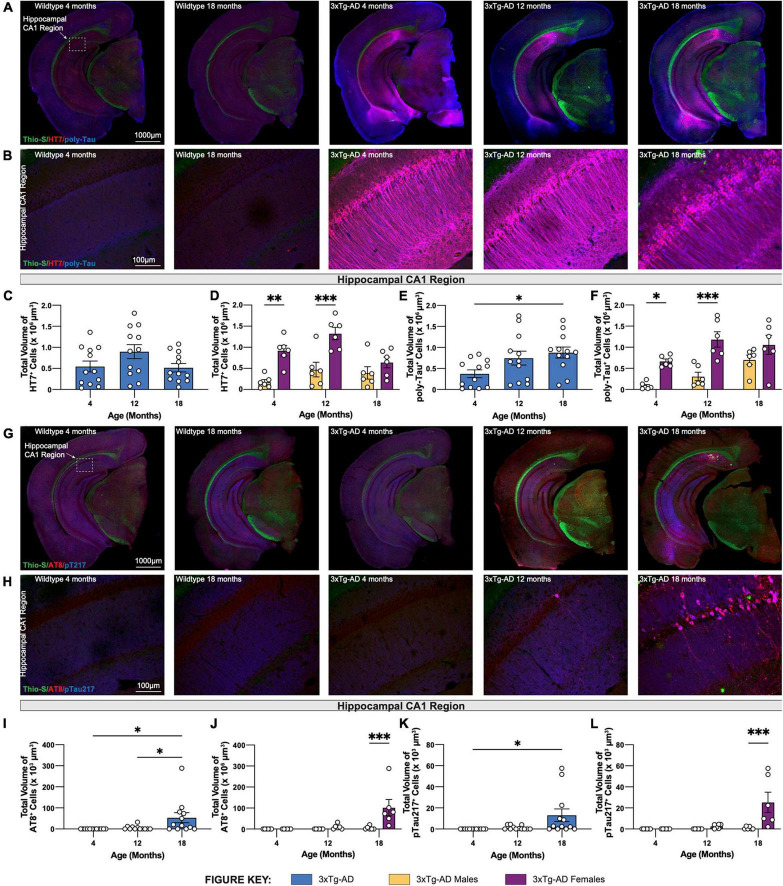
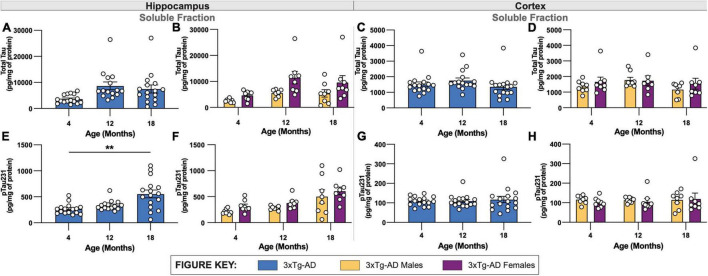
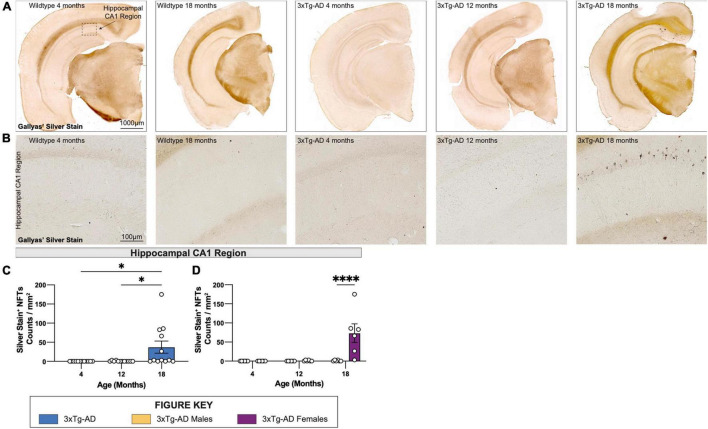
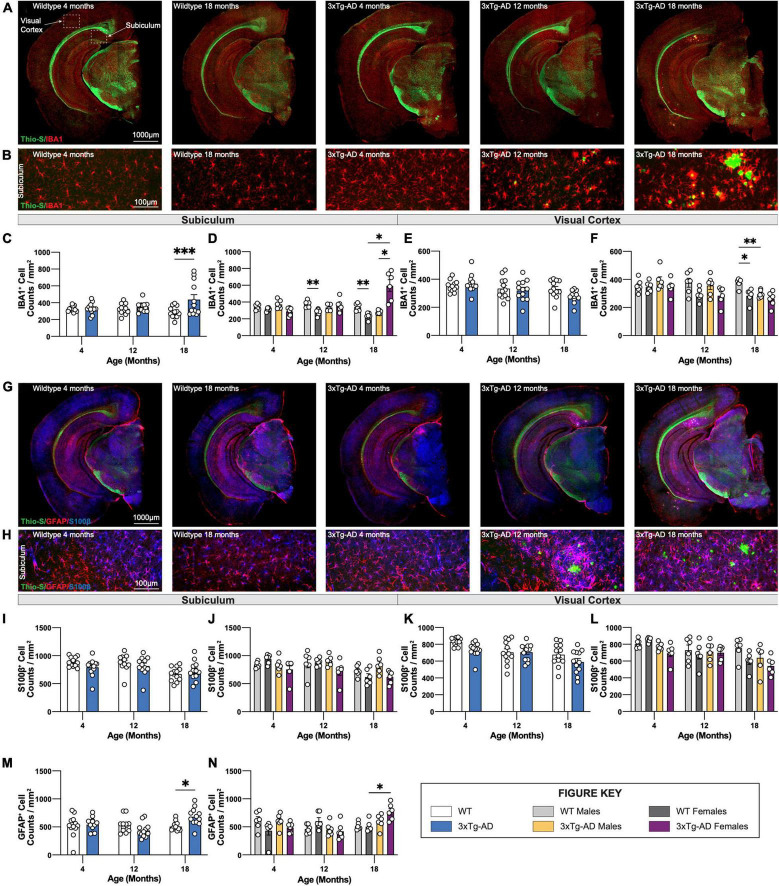
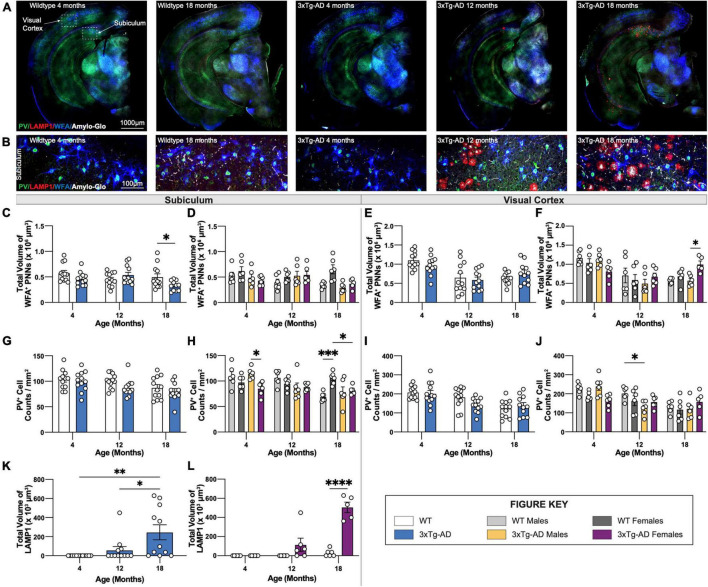
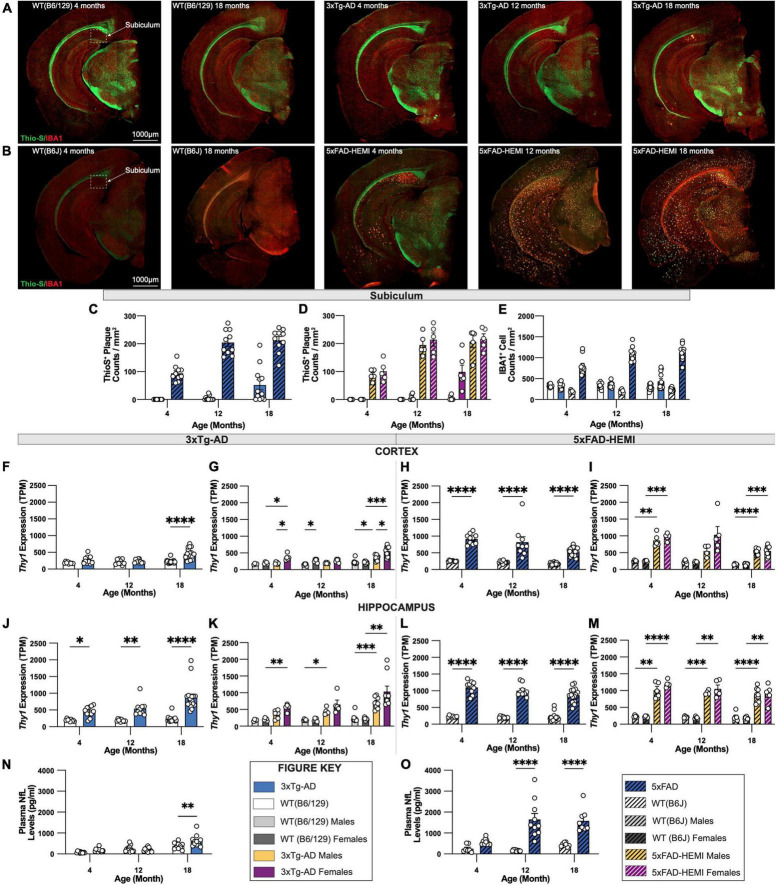
Animal models of disease are valuable resources for investigating pathogenic mechanisms and potential therapeutic interventions. However, for complex disorders such as Alzheimer's disease (AD), the generation and availability of innumerous distinct animal models present unique challenges to AD researchers and hinder the success of useful therapies. Here, we conducted an in-depth analysis of the 3xTg-AD mouse model of AD across its lifespan to better inform the field of the various pathologies that appear at specific ages, and comment on drift that has occurred in the development of pathology in this line since its development 20 years ago. This modern characterization of the 3xTg-AD model includes an assessment of impairments in long-term potentiation followed by quantification of amyloid beta (Aβ) plaque burden and neurofibrillary tau tangles, biochemical levels of Aβ and tau protein, and neuropathological markers such as gliosis and accumulation of dystrophic neurites. We also present a novel comparison of the 3xTg-AD model with the 5xFAD model using the same deep-phenotyping characterization pipeline and show plasma NfL is strongly driven by plaque burden. The results from these analyses are freely available via the AD Knowledge Portal (https://modeladexplorer.org/). Our work demonstrates the utility of a characterization pipeline that generates robust and standardized information relevant to investigating and comparing disease etiologies of current and future models of AD.
Keywords: 3xTg-AD; Alzheimer’s disease; amyloid precursor protein; amyloid β-protein; animal model; genetically modified; neurofibrillary tangles; tau.
Copyright © 2022 Javonillo, Tran, Phan, Hingco, Kramár, da Cunha, Forner, Kawauchi, Milinkeviciute, Gomez-Arboledas, Neumann, Banh, Huynh, Matheos, Rezaie, Alcantara, Mortazavi, Wood, Tenner, MacGregor, Green and LaFerla.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Balderrama-Gutierrez G., Liang H., Rezaie N., Carvalho K., Forner S., Matheos D., et al. (2021). Single-cell and nucleus RNA-seq in a mouse model of AD reveal activation of distinct glial subpopulations in the presence of plaques and tangles. bioRxiv [preprint]. 10.1101/2021.09.29.462436 - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases