

Exercise improves angiogenic function of circulating exosomes in type 2 diabetes: Role of exosomal SOD3
- PMID: 35142393
- PMCID: PMC8880294
- DOI: 10.1096/fj.202101323R
Exercise improves angiogenic function of circulating exosomes in type 2 diabetes: Role of exosomal SOD3
Abstract
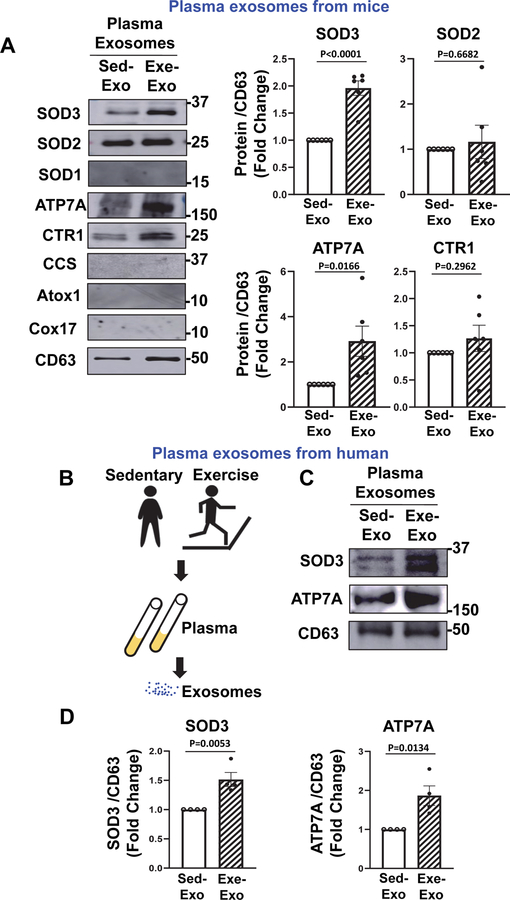
Exosomes, key mediators of cell-cell communication, derived from type 2 diabetes mellitus (T2DM) exhibit detrimental effects. Exercise improves endothelial function in part via the secretion of exosomes into circulation. Extracellular superoxide dismutase (SOD3) is a major secretory copper (Cu) antioxidant enzyme that catalyzes the dismutation of O2•- to H2 O2 whose activity requires the Cu transporter ATP7A. However, the role of SOD3 in exercise-induced angiogenic effects of circulating plasma exosomes on endothelial cells (ECs) in T2DM remains unknown. Here, we show that both SOD3 and ATP7A proteins were present in plasma exosomes in mice, which was significantly increased after two weeks of volunteer wheel exercise. A single bout of exercise in humans also showed a significant increase in SOD3 and ATP7A protein expression in plasma exosomes. Plasma exosomes from T2DM mice significantly reduced angiogenic responses in human ECs or mouse skin wound healing models, which was associated with a decrease in ATP7A, but not SOD3 expression in exosomes. Exercise training in T2DM mice restored the angiogenic effects of T2DM exosomes in ECs by increasing ATP7A in exosomes, which was not observed in exercised T2DM/SOD3-/- mice. Furthermore, exosomes overexpressing SOD3 significantly enhanced angiogenesis in ECs by increasing local H2 O2 levels in a heparin-binding domain-dependent manner as well as restored defective wound healing and angiogenesis in T2DM or SOD3-/- mice. In conclusion, exercise improves the angiogenic potential of circulating exosomes in T2DM in a SOD3-dependent manner. Exosomal SOD3 may provide an exercise mimetic therapy that supports neovascularization and wound repair in cardiometabolic disease.
Keywords: SOD3; exercise; exosome; type 2 diabetes.
© 2022 Federation of American Societies for Experimental Biology.
Conflict of interest statement
DISCLOSURES
The authors declare no competing financial interests.
Figures







References
-
- Tkach M, and Thery C (2016) Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 164, 1226–1232 - PubMed
-
- Lawson C, Vicencio JM, Yellon DM, and Davidson SM (2016) Microvesicles and exosomes: new players in metabolic and cardiovascular disease. J Endocrinol 228, R57–71 - PubMed
-
- Kranendonk ME, de Kleijn DP, Kalkhoven E, Kanhai DA, Uiterwaal CS, van der Graaf Y, Pasterkamp G, Visseren FL, and Group SS (2014) Extracellular vesicle markers in relation to obesity and metabolic complications in patients with manifest cardiovascular disease. Cardiovasc Diabetol 13, 37. - PMC - PubMed
-
- Yanez-Mo M, Siljander PR, Andreu Z, Zavec AB, Borras FE, Buzas EI, Buzas K, Casal E, Cappello F, Carvalho J, Colas E, Cordeiro-da Silva A, Fais S, Falcon-Perez JM, Ghobrial IM, Giebel B, Gimona M, Graner M, Gursel I, Gursel M, Heegaard NH, Hendrix A, Kierulf P, Kokubun K, Kosanovic M, Kralj-Iglic V, Kramer-Albers EM, Laitinen S, Lasser C, Lener T, Ligeti E, Line A, Lipps G, Llorente A, Lotvall J, Mancek-Keber M, Marcilla A, Mittelbrunn M, Nazarenko I, Nolte-’t Hoen EN, Nyman TA, O’Driscoll L, Olivan M, Oliveira C, Pallinger E, Del Portillo HA, Reventos J, Rigau M, Rohde E, Sammar M, Sanchez-Madrid F, Santarem N, Schallmoser K, Ostenfeld MS, Stoorvogel W, Stukelj R, Van der Grein SG, Vasconcelos MH, Wauben MH, and De Wever O (2015) Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles 4, 27066. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
