

Caloric restriction in humans reveals immunometabolic regulators of health span
- PMID: 35143297
- PMCID: PMC10061495
- DOI: 10.1126/science.abg7292
Caloric restriction in humans reveals immunometabolic regulators of health span
Abstract
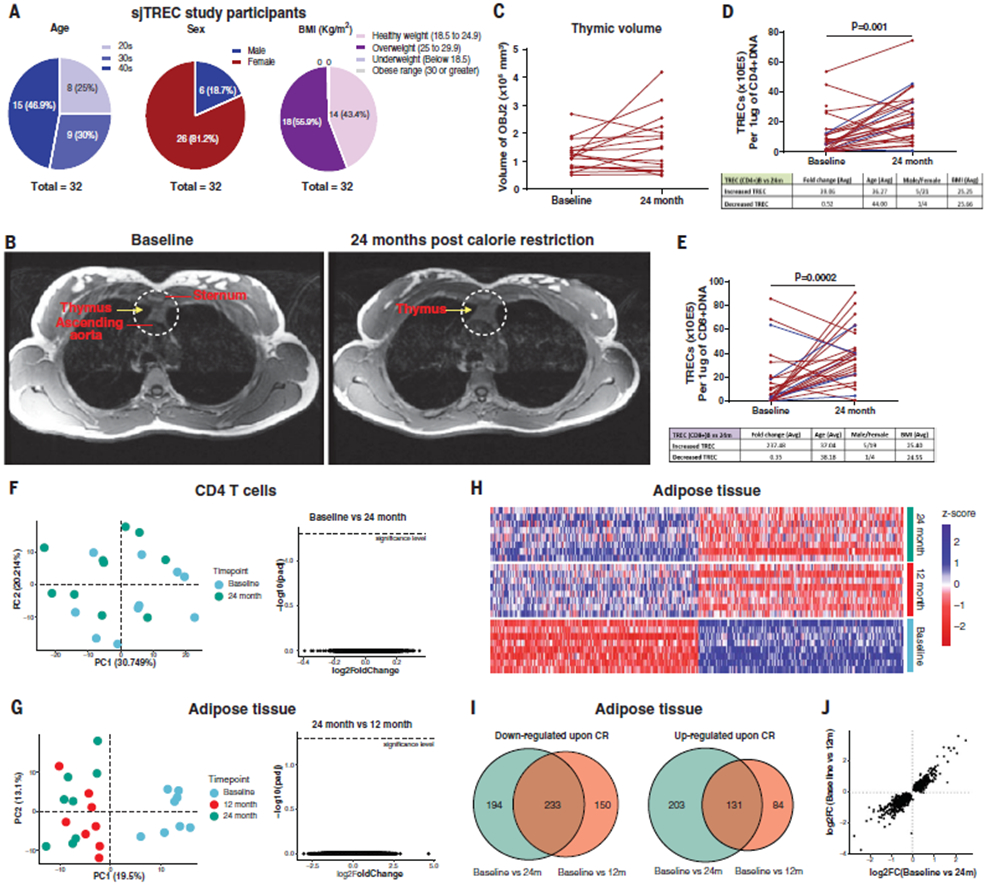
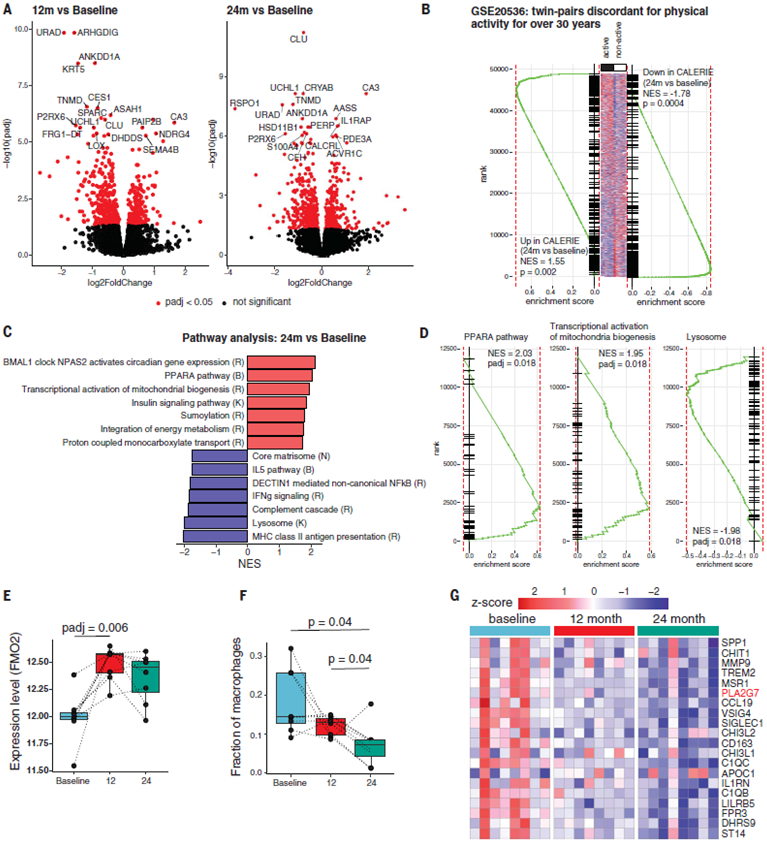
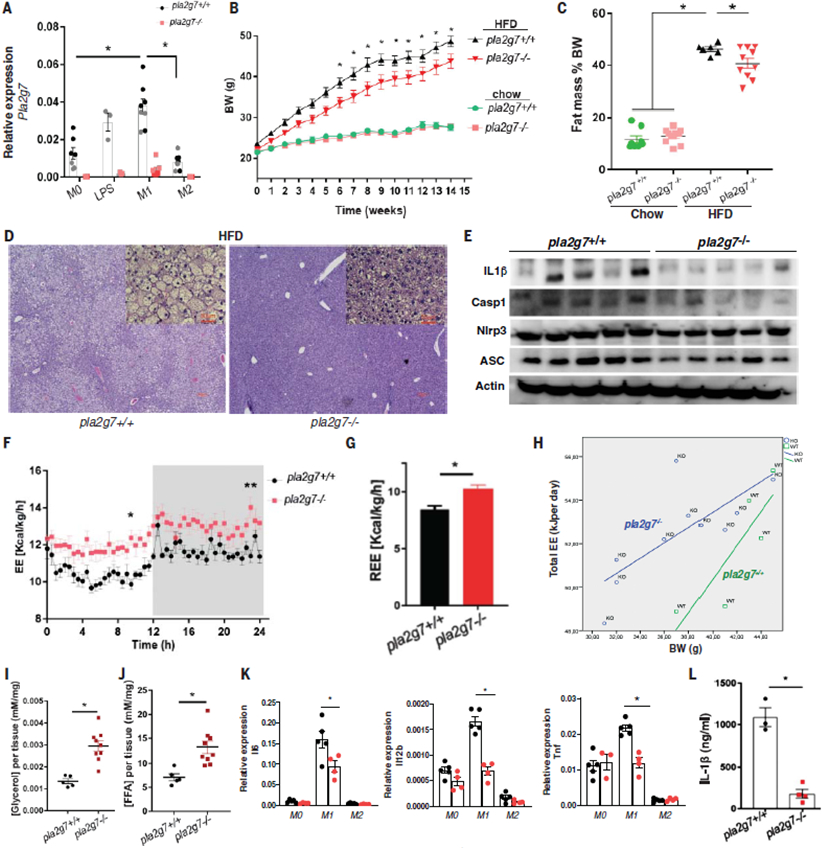
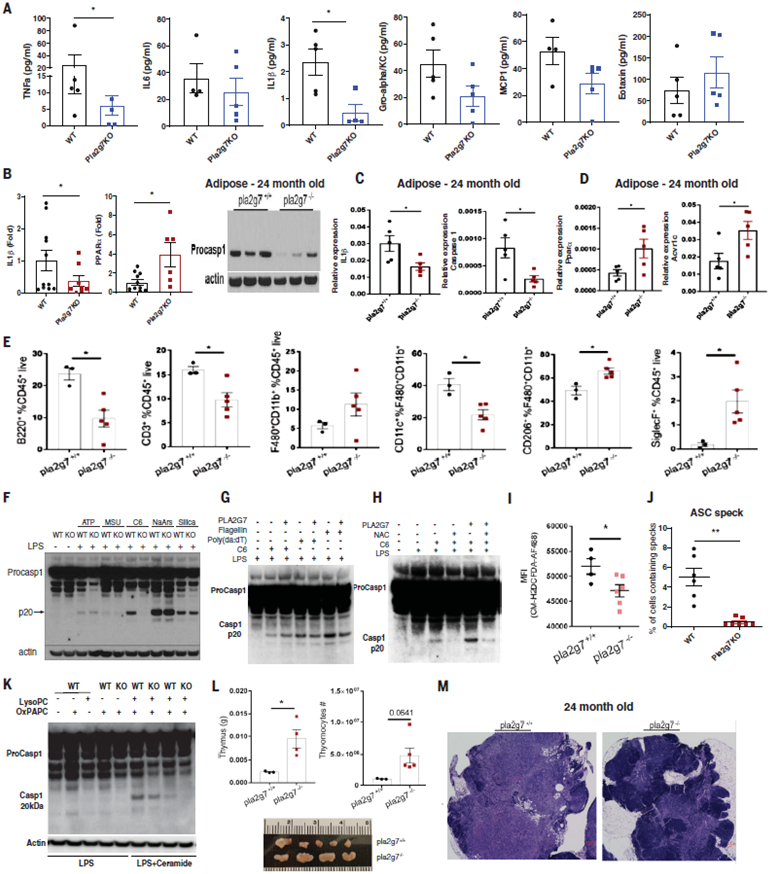
The extension of life span driven by 40% caloric restriction (CR) in rodents causes trade-offs in growth, reproduction, and immune defense that make it difficult to identify therapeutically relevant CR-mimetic targets. We report that about 14% CR for 2 years in healthy humans improved thymopoiesis and was correlated with mobilization of intrathymic ectopic lipid. CR-induced transcriptional reprogramming in adipose tissue implicated pathways regulating mitochondrial bioenergetics, anti-inflammatory responses, and longevity. Expression of the gene Pla2g7 encoding platelet activating factor acetyl hydrolase (PLA2G7) is inhibited in humans undergoing CR. Deletion of Pla2g7 in mice showed decreased thymic lipoatrophy, protection against age-related inflammation, lowered NLRP3 inflammasome activation, and improved metabolic health. Therefore, the reduction of PLA2G7 may mediate the immunometabolic effects of CR and could potentially be harnessed to lower inflammation and extend the health span.
Figures
Comment in
-
Caloric restriction has a new player.Science. 2022 Feb 11;375(6581):620-621. doi: 10.1126/science.abn6576. Epub 2022 Feb 10. Science. 2022. PMID: 35143311
-
PLA2G7, caloric restriction and cardiovascular aging.J Cardiovasc Aging. 2022 Apr;2(2):19. doi: 10.20517/jca.2022.08. Epub 2022 Mar 16. J Cardiovasc Aging. 2022. PMID: 35497092 Free PMC article. No abstract available.
-
PLA2G7: a new player in shaping energy metabolism and lifespan.Signal Transduct Target Ther. 2022 Jun 17;7(1):195. doi: 10.1038/s41392-022-01052-5. Signal Transduct Target Ther. 2022. PMID: 35715398 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
