

Comparative Genomics Elucidates the Origin of a Supergene Controlling Floral Heteromorphism
- PMID: 35143659
- PMCID: PMC8859637
- DOI: 10.1093/molbev/msac035
Comparative Genomics Elucidates the Origin of a Supergene Controlling Floral Heteromorphism
Abstract
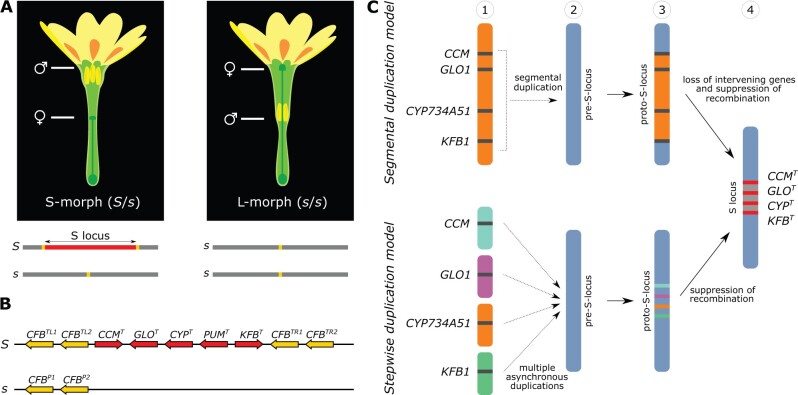
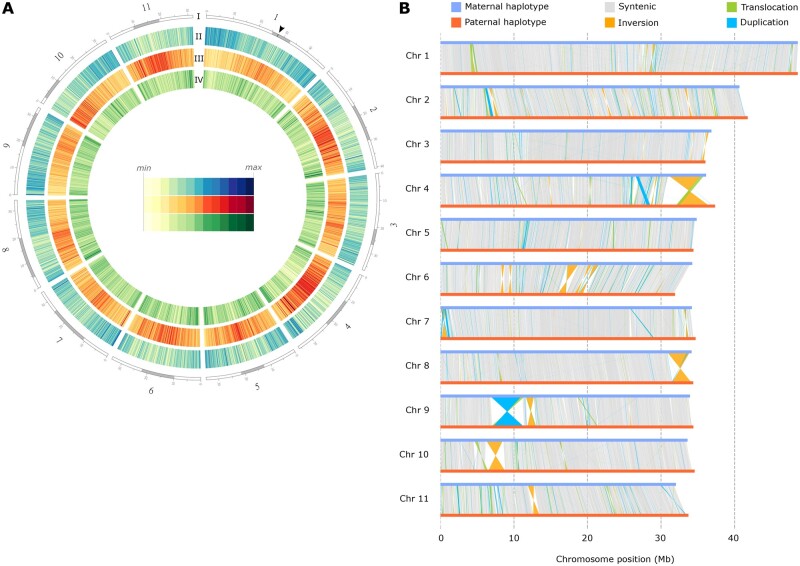
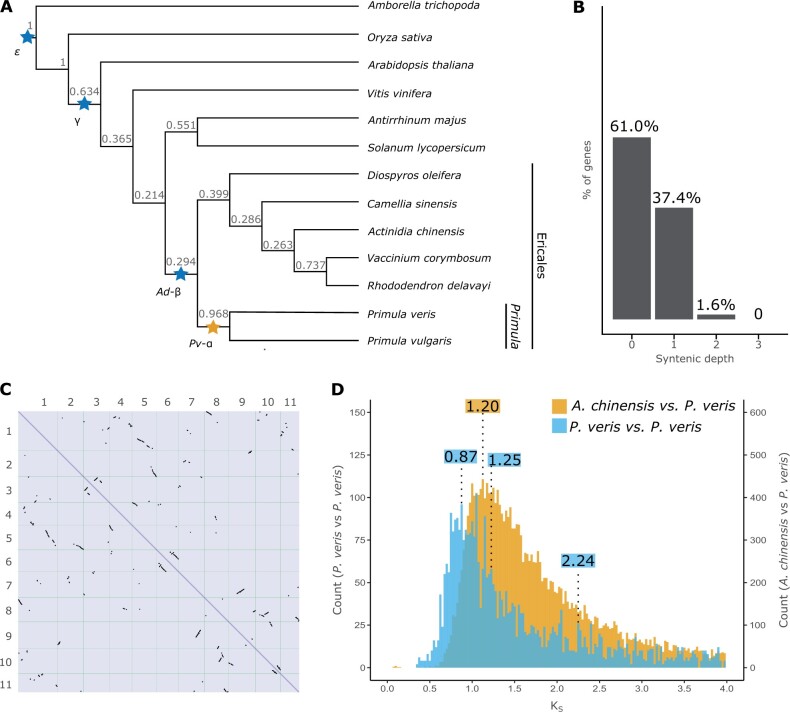
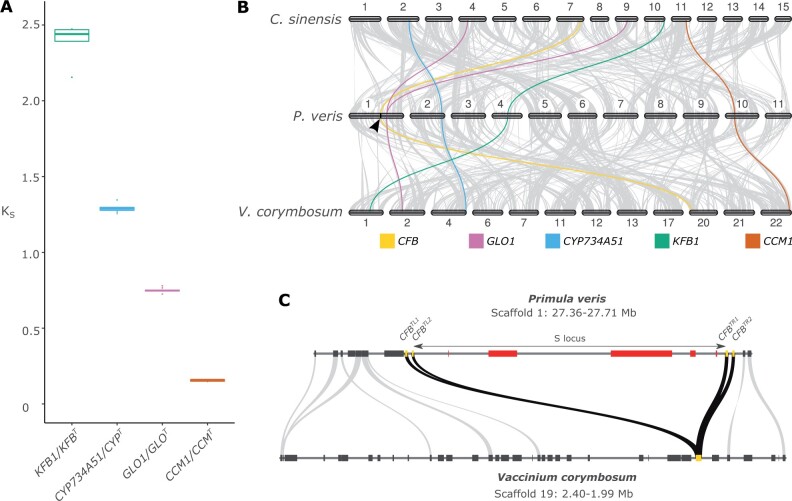
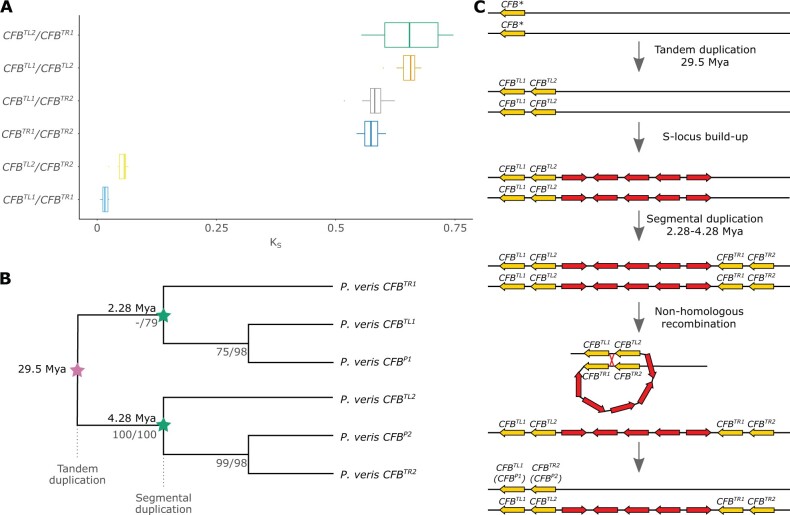
Supergenes are nonrecombining genomic regions ensuring the coinheritance of multiple, coadapted genes. Despite the importance of supergenes in adaptation, little is known on how they originate. A classic example of supergene is the S locus controlling heterostyly, a floral heteromorphism occurring in 28 angiosperm families. In Primula, heterostyly is characterized by the cooccurrence of two complementary, self-incompatible floral morphs and is controlled by five genes clustered in the hemizygous, ca. 300-kb S locus. Here, we present the first chromosome-scale genome assembly of any heterostylous species, that of Primula veris (cowslip). By leveraging the high contiguity of the P. veris assembly and comparative genomic analyses, we demonstrated that the S-locus evolved via multiple, asynchronous gene duplications and independent gene translocations. Furthermore, we discovered a new whole-genome duplication in Ericales that is specific to the Primula lineage. We also propose a mechanism for the origin of S-locus hemizygosity via nonhomologous recombination involving the newly discovered two pairs of CFB genes flanking the S locus. Finally, we detected only weak signatures of degeneration in the S locus, as predicted for hemizygous supergenes. The present study provides a useful resource for future research addressing key questions on the evolution of supergenes in general and the S locus in particular: How do supergenes arise? What is the role of genome architecture in the evolution of complex adaptations? Is the molecular architecture of heterostyly supergenes across angiosperms similar to that of Primula?
Keywords: chromosome-scale genome assembly; evolutionary genomics; genome architecture; heterostyly; primula; supergene.
© The Author(s) 2022. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Barrett SCH. 2002. The evolution of plant sexual diversity. Nat Rev Genet. 3(4):274–284. - PubMed
-
- Barrett SCH. 2019. ‘A most complex marriage arrangement’: recent advances on heterostyly and unresolved questions. New Phytol. 224(3):1051–1067. - PubMed
-
- Becher H, Jackson BC, Charlesworth B.. 2020. Patterns of genetic variability in genomic regions with low rates of recombination. Curr Biol. 30(1):94–100.e3. - PubMed
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
