

dCas9-based gene editing for cleavage-free genomic knock-in of long sequences
- PMID: 35145221
- PMCID: PMC8843813
- DOI: 10.1038/s41556-021-00836-1
dCas9-based gene editing for cleavage-free genomic knock-in of long sequences
Abstract
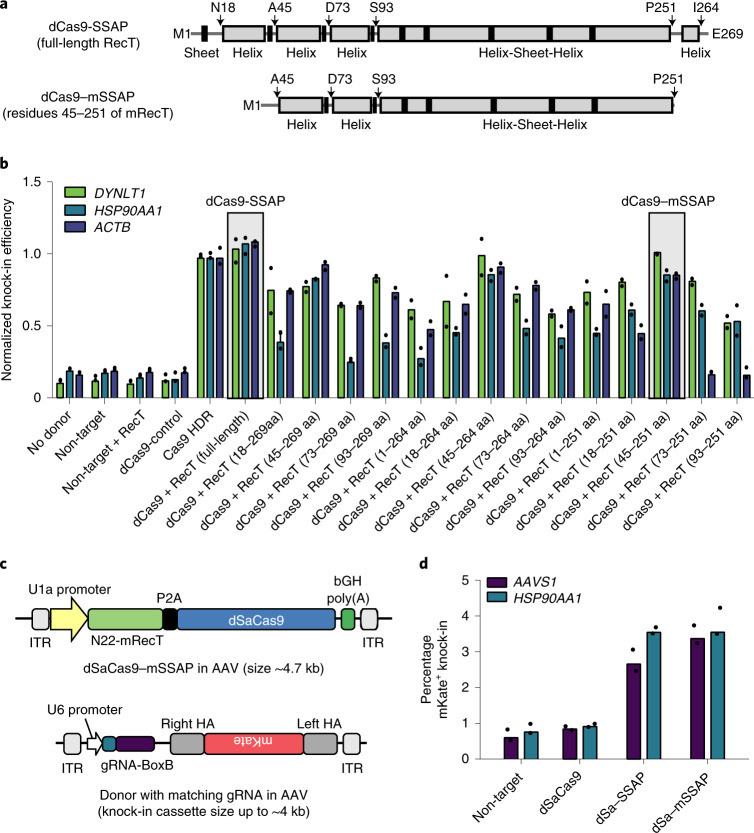
Gene editing is a powerful tool for genome and cell engineering. Exemplified by CRISPR-Cas, gene editing could cause DNA damage and trigger DNA repair processes that are often error-prone. Such unwanted mutations and safety concerns can be exacerbated when altering long sequences. Here we couple microbial single-strand annealing proteins (SSAPs) with catalytically inactive dCas9 for gene editing. This cleavage-free gene editor, dCas9-SSAP, promotes the knock-in of long sequences in mammalian cells. The dCas9-SSAP editor has low on-target errors and minimal off-target effects, showing higher accuracy than canonical Cas9 methods. It is effective for inserting kilobase-scale sequences, with an efficiency of up to approximately 20% and robust performance across donor designs and cell types, including human stem cells. We show that dCas9-SSAP is less sensitive to inhibition of DNA repair enzymes than Cas9 references. We further performed truncation and aptamer engineering to minimize its size to fit into a single adeno-associated-virus vector for future application. Together, this tool opens opportunities towards safer long-sequence genome engineering.
© 2022. The Author(s).
Conflict of interest statement
Stanford University has filed patent applications with L.C., C.W. and Y.Q. as inventors based on this work.
Figures

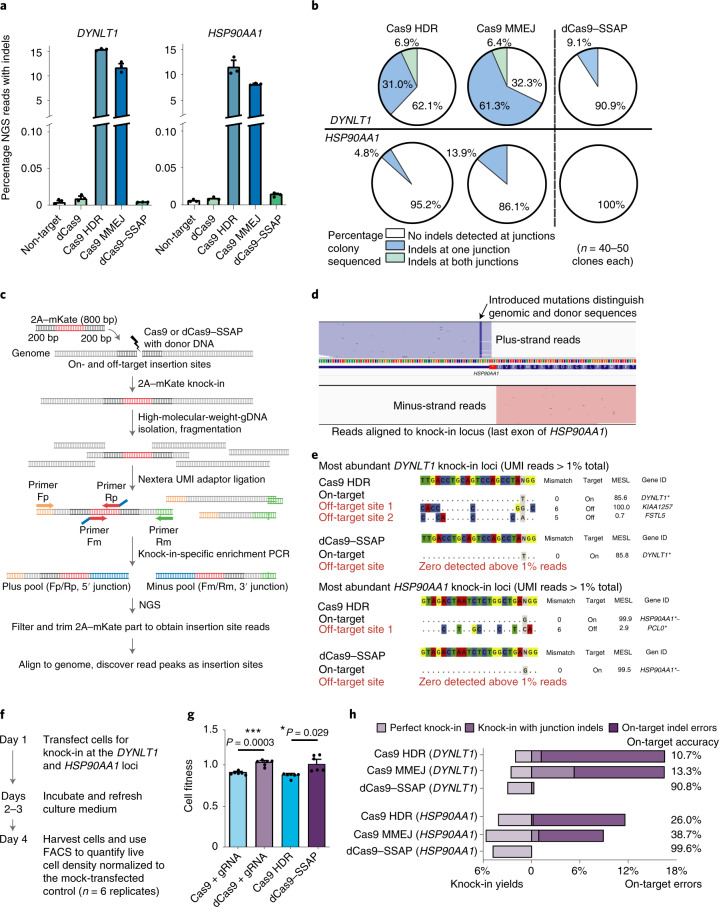
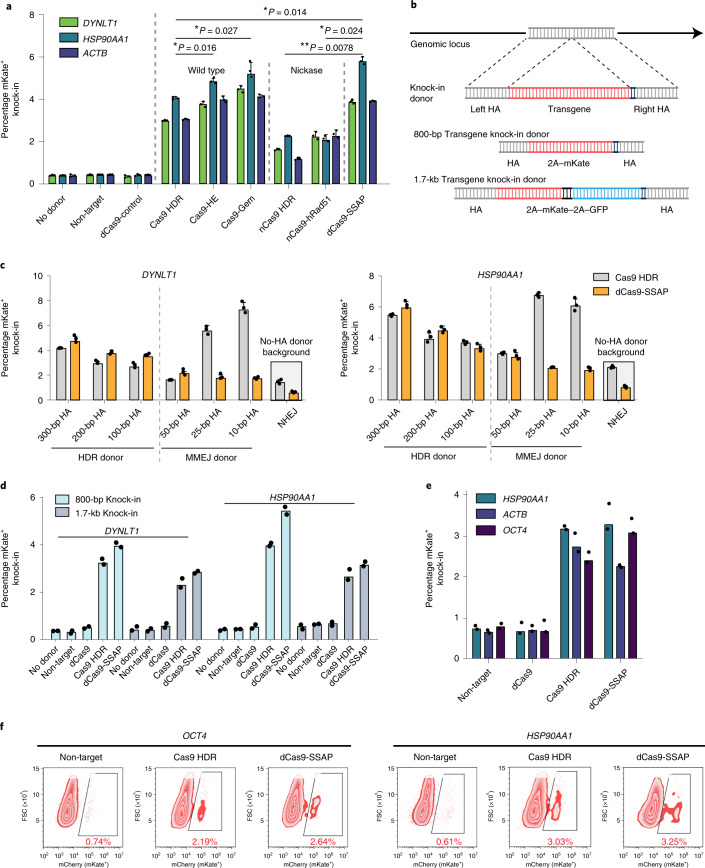
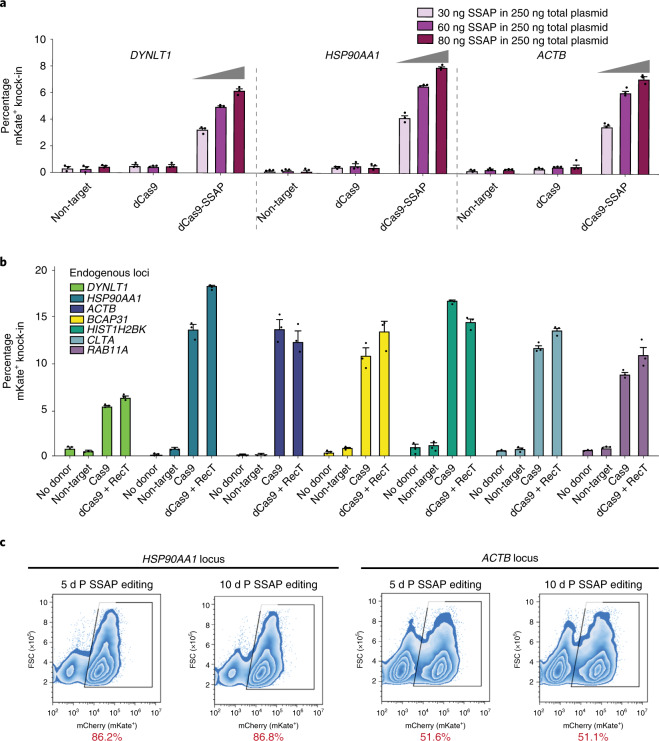
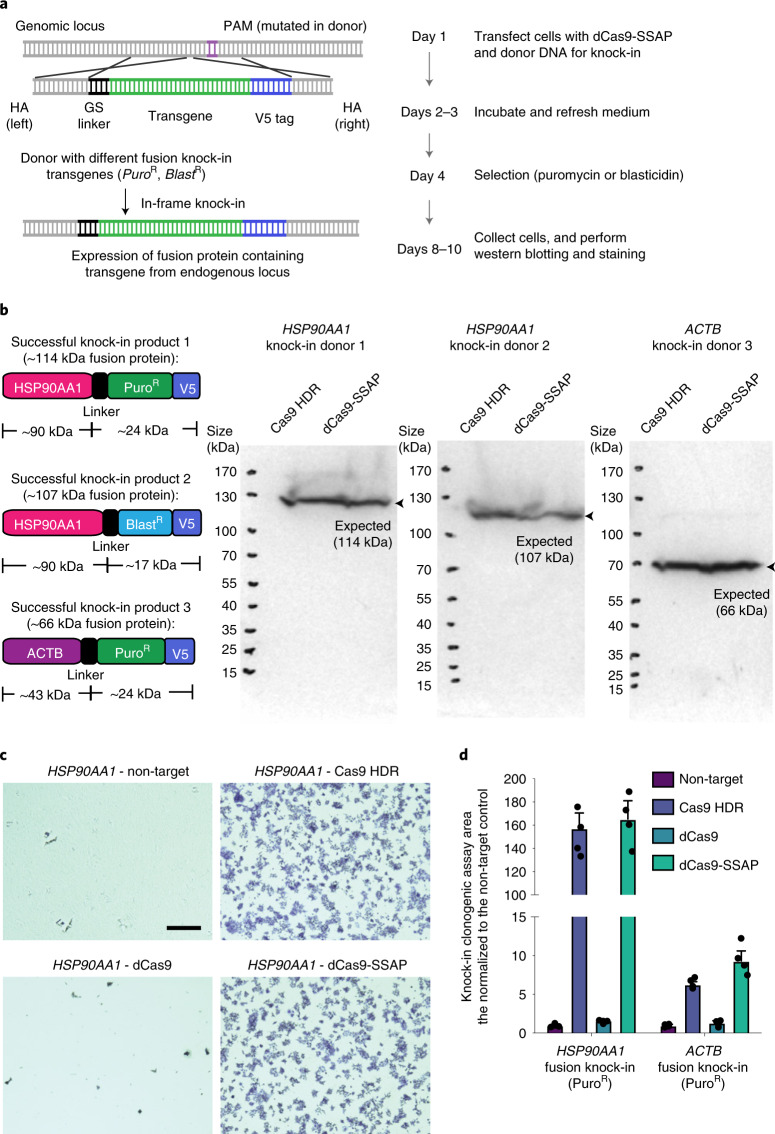











References
-
- Carroll D. Genome engineering with targetable nucleases. Annu. Rev. Biochem. 2014;83:409–439. - PubMed
-
- Barrangou R, Doudna JA. Applications of CRISPR technologies in research and beyond. Nat. Biotechnol. 2016;34:933–941. - PubMed
-
- Doudna JA, Charpentier E. The new frontier of genome engineering with CRISPR–Cas9. Science. 2014;346:1258096. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
