

Efficient in vitro and in vivo RNA editing via recruitment of endogenous ADARs using circular guide RNAs
- PMID: 35145312
- PMCID: PMC9232839
- DOI: 10.1038/s41587-021-01171-4
Efficient in vitro and in vivo RNA editing via recruitment of endogenous ADARs using circular guide RNAs
Abstract
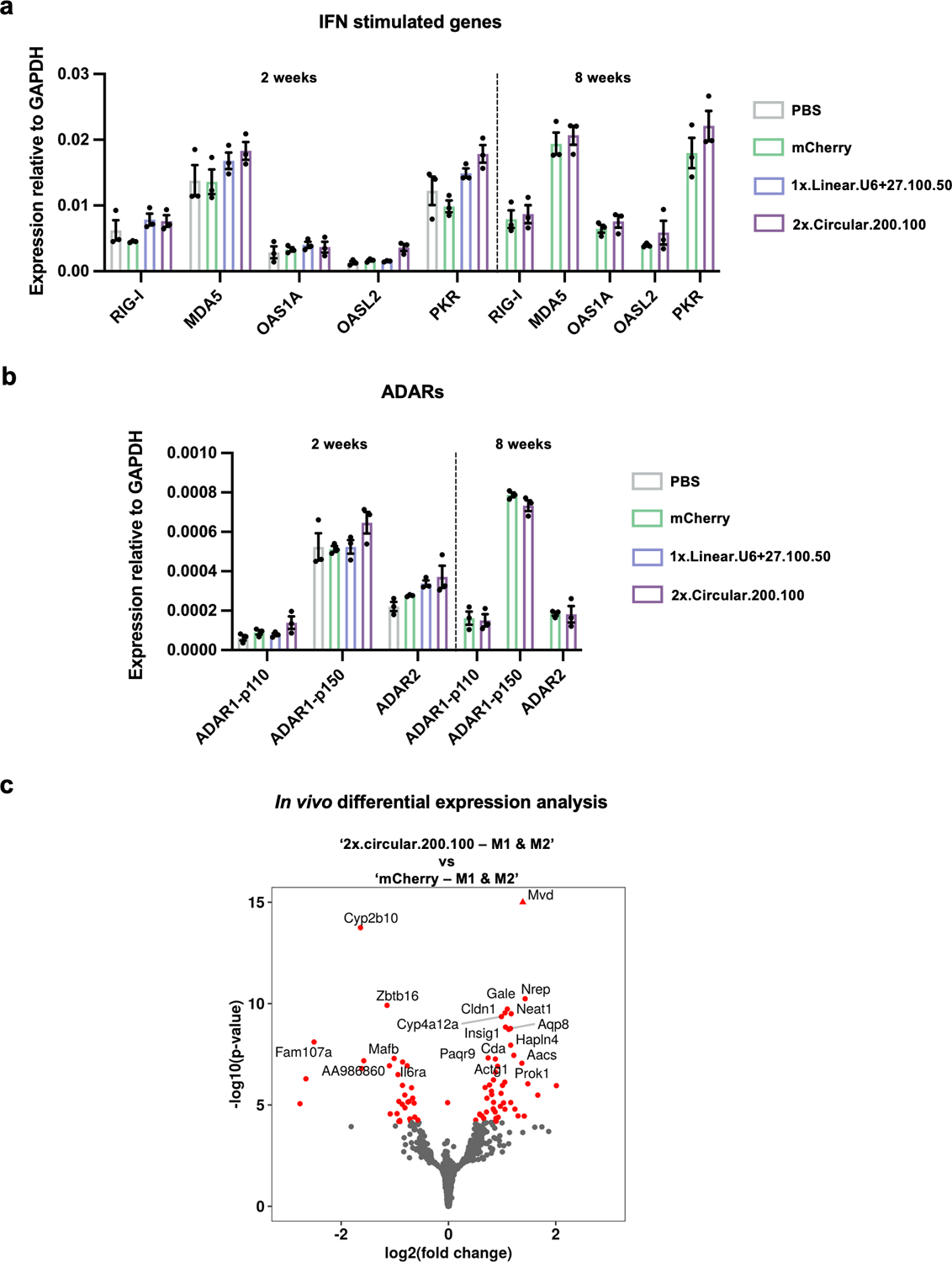
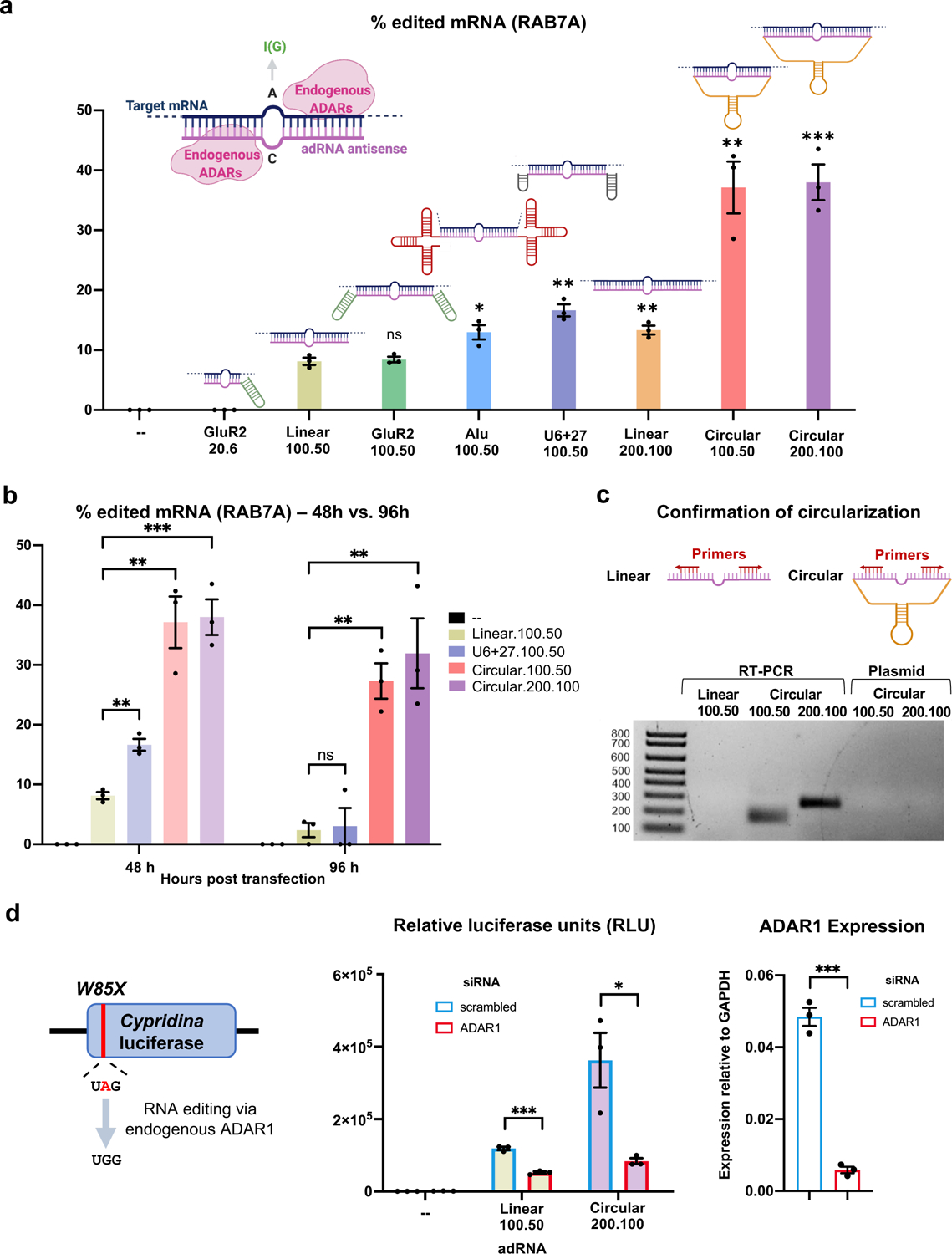
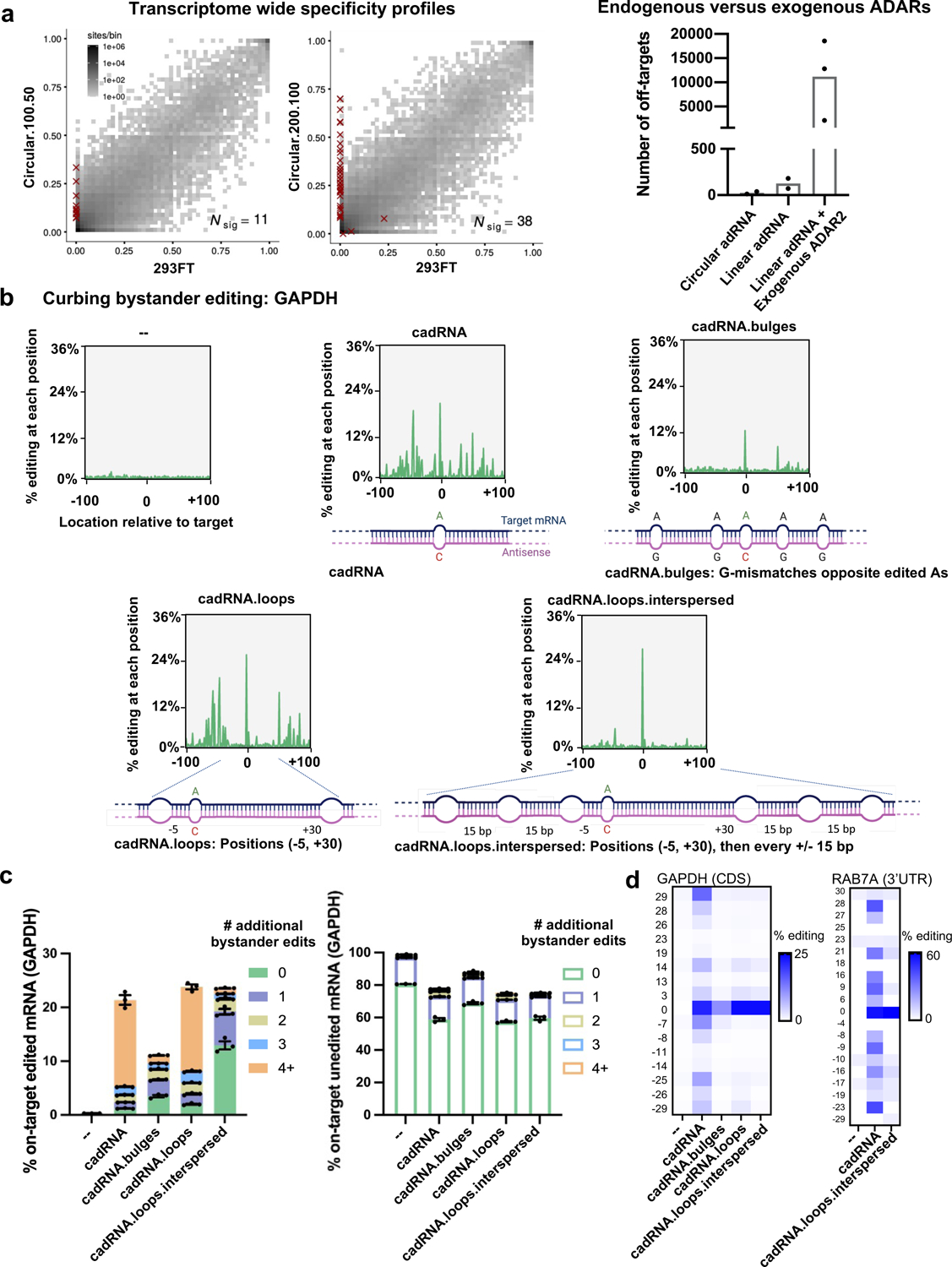
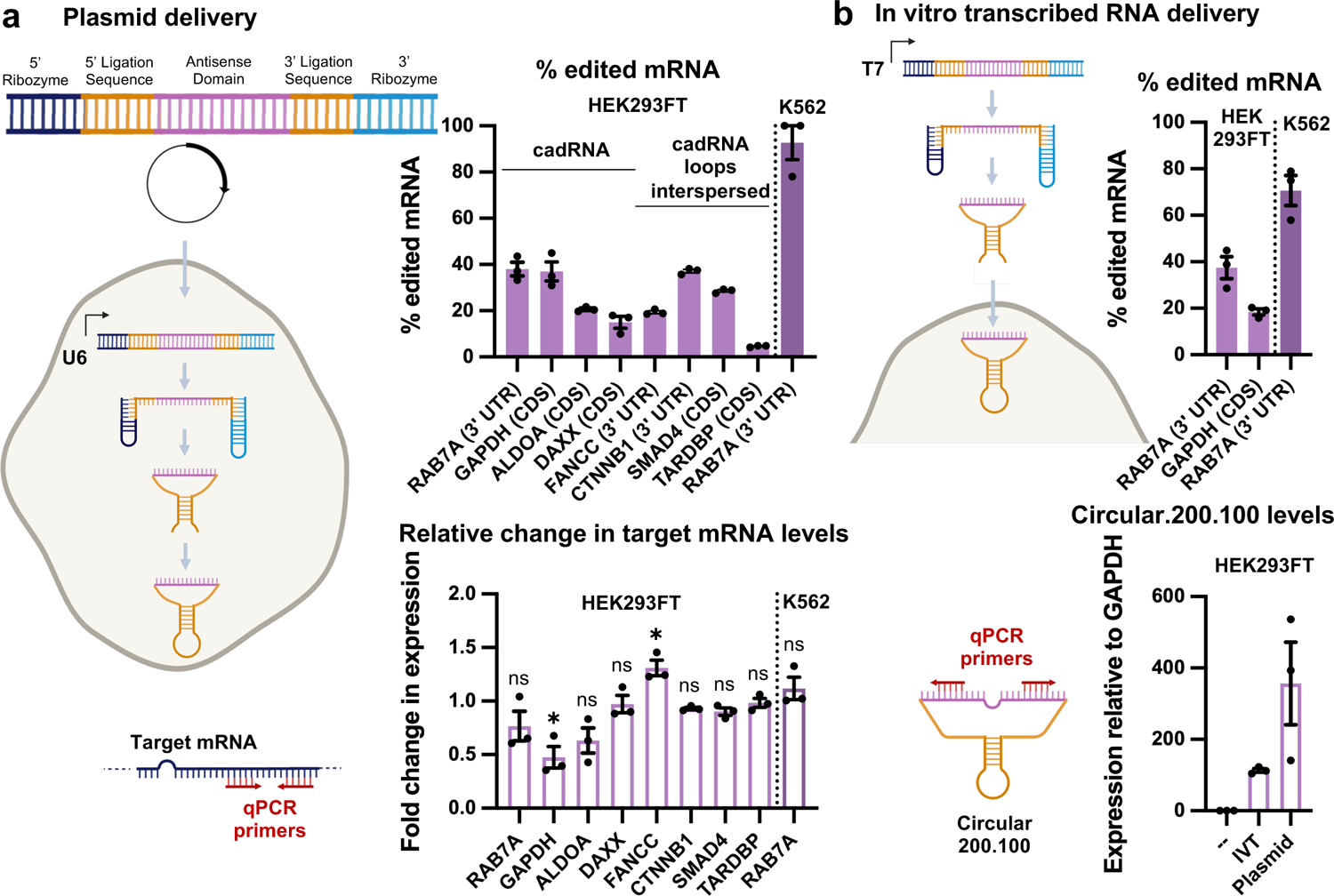
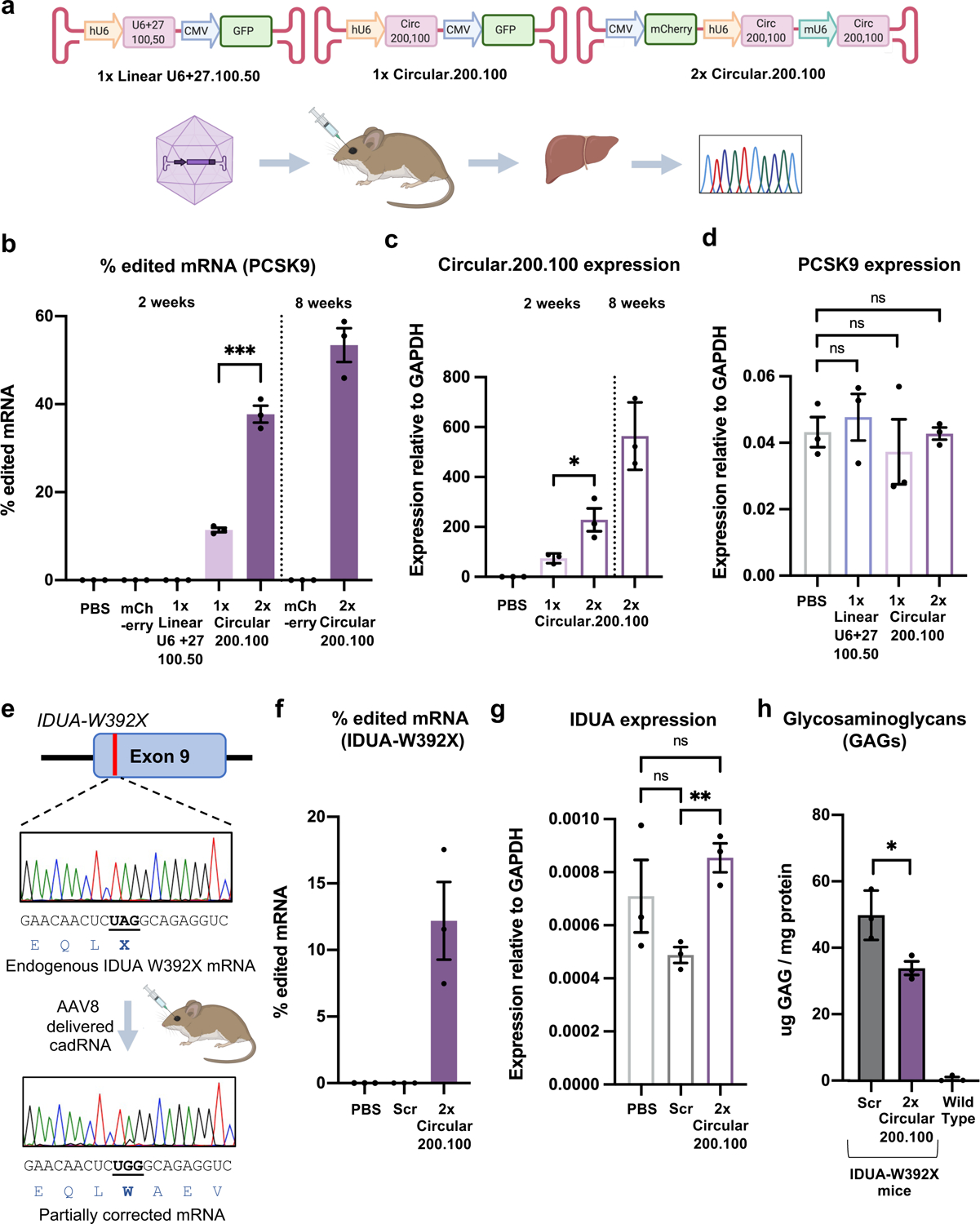
Recruiting endogenous adenosine deaminases using exogenous guide RNAs to edit cellular RNAs is a promising therapeutic strategy, but editing efficiency and durability remain low using current guide RNA designs. In this study, we engineered circular ADAR-recruiting guide RNAs (cadRNAs) to enable more efficient programmable adenosine-to-inosine RNA editing without requiring co-delivery of any exogenous proteins. Using these cadRNAs, we observed robust and durable RNA editing across multiple sites and cell lines, in both untranslated and coding regions of RNAs, and high transcriptome-wide specificity. Additionally, we increased transcript-level specificity for the target adenosine by incorporating interspersed loops in the antisense domains, reducing bystander editing. In vivo delivery of cadRNAs via adeno-associated viruses enabled 53% RNA editing of the mPCSK9 transcript in C57BL/6J mice livers and 12% UAG-to-UGG RNA correction of the amber nonsense mutation in the IDUA-W392X mouse model of mucopolysaccharidosis type I-Hurler syndrome. cadRNAs enable efficient programmable RNA editing in vivo with diverse protein modulation and gene therapeutic applications.
© 2022. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
COMPETING FINANCIAL INTERESTS
D.K. and P.M. have filed patents based on this work. P.M. is a scientific co-founder of Shape Therapeutics, Boundless Biosciences, Seven Therapeutics, Navega Therapeutics, and Engine Biosciences. The terms of these arrangements have been reviewed and approved by the University of California, San Diego in accordance with its conflict of interest policies. Y.S. is an employee of Shape Therapeutics. D.K. is now an employee of Shape Therapeutics. The remaining authors declare no competing interests.
Figures
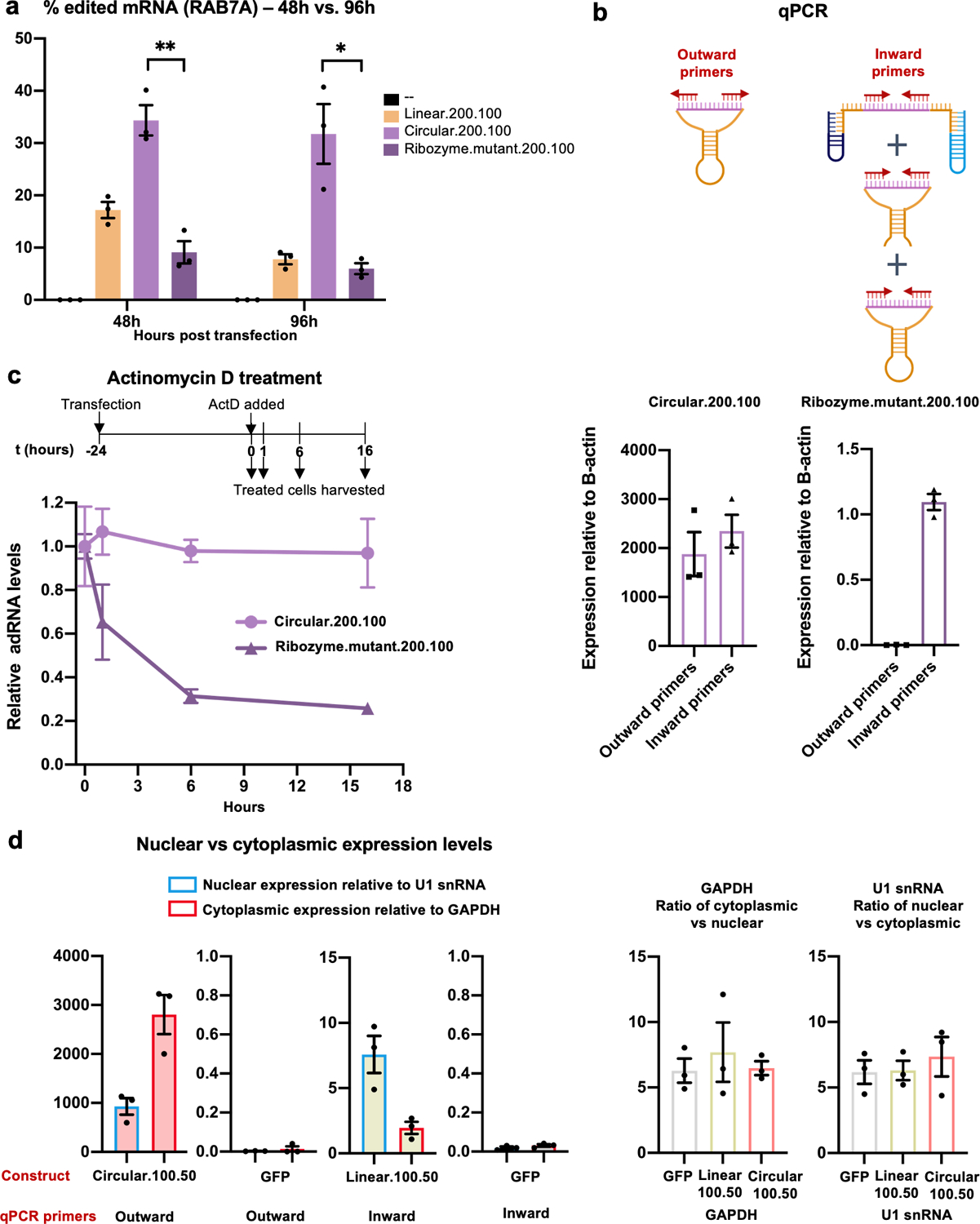
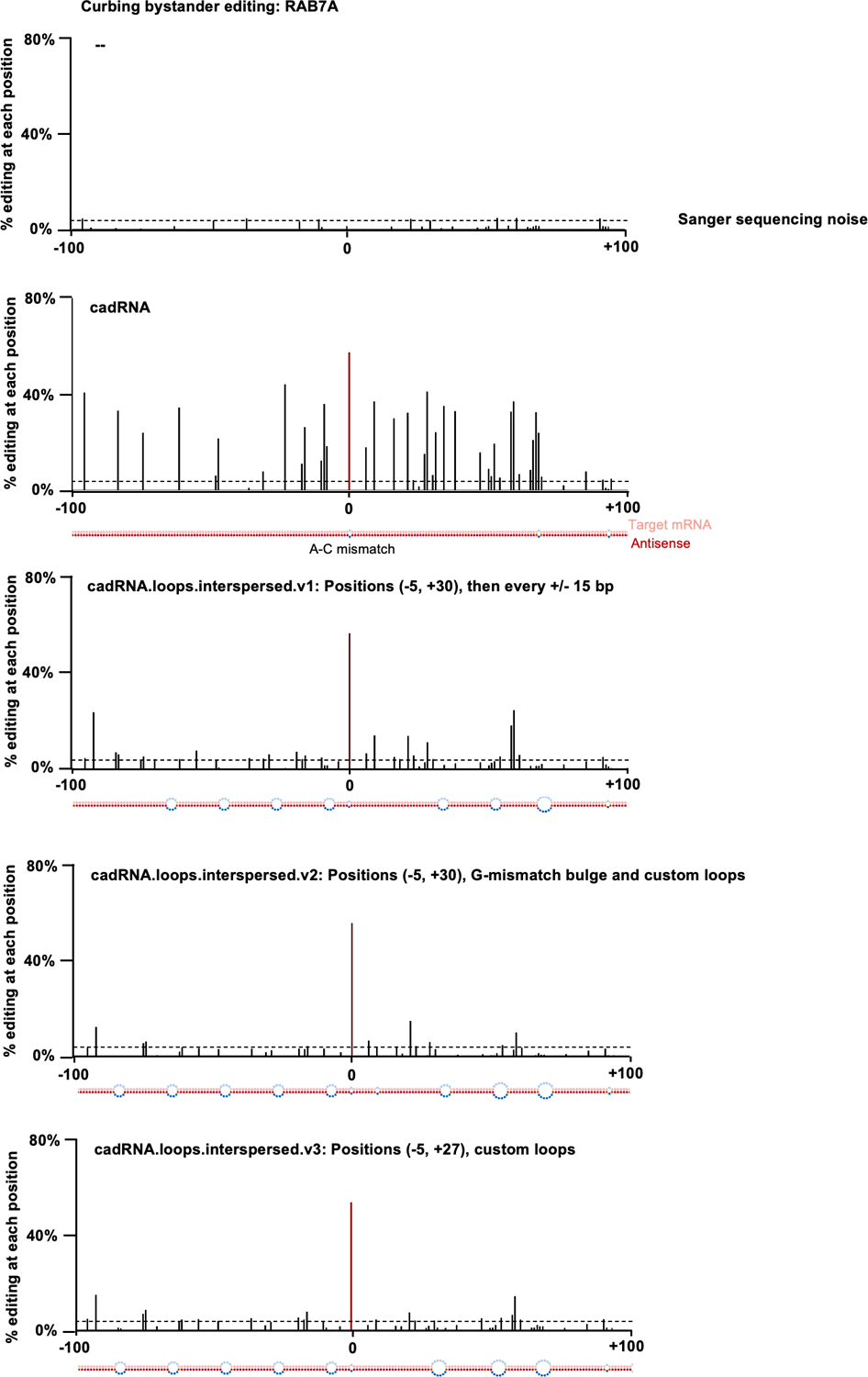
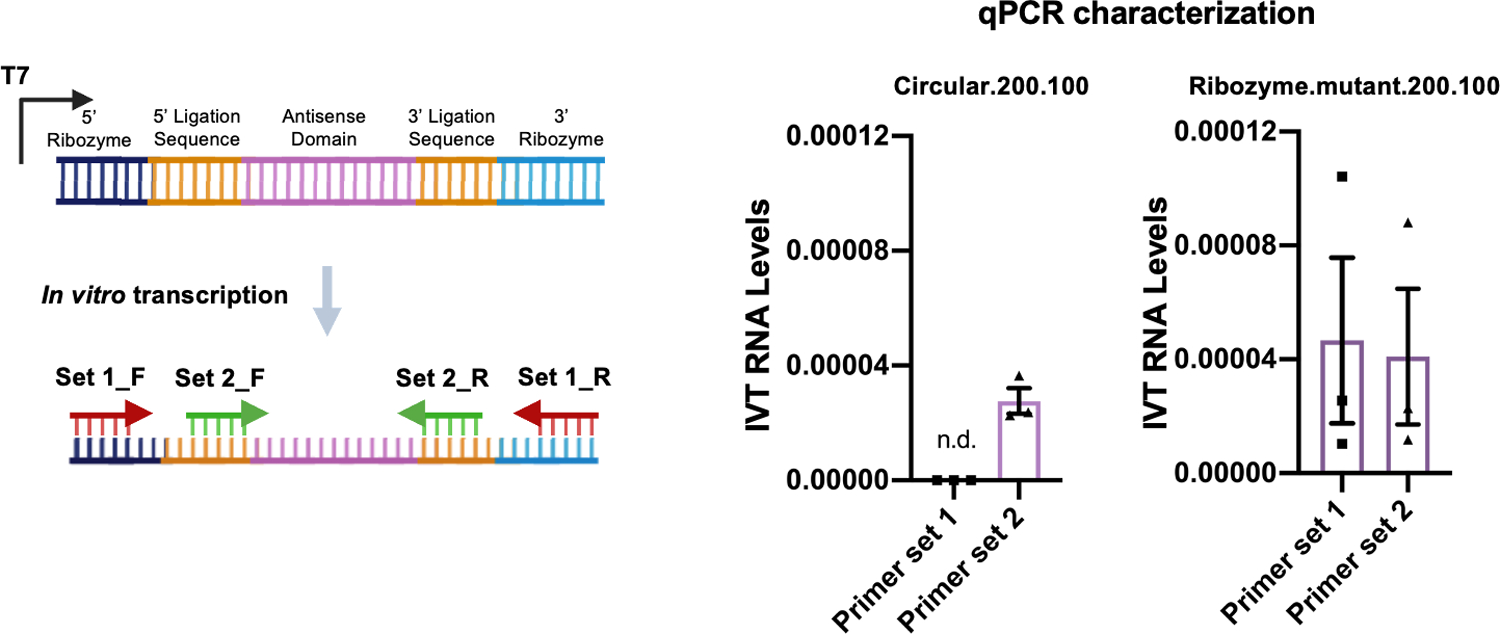
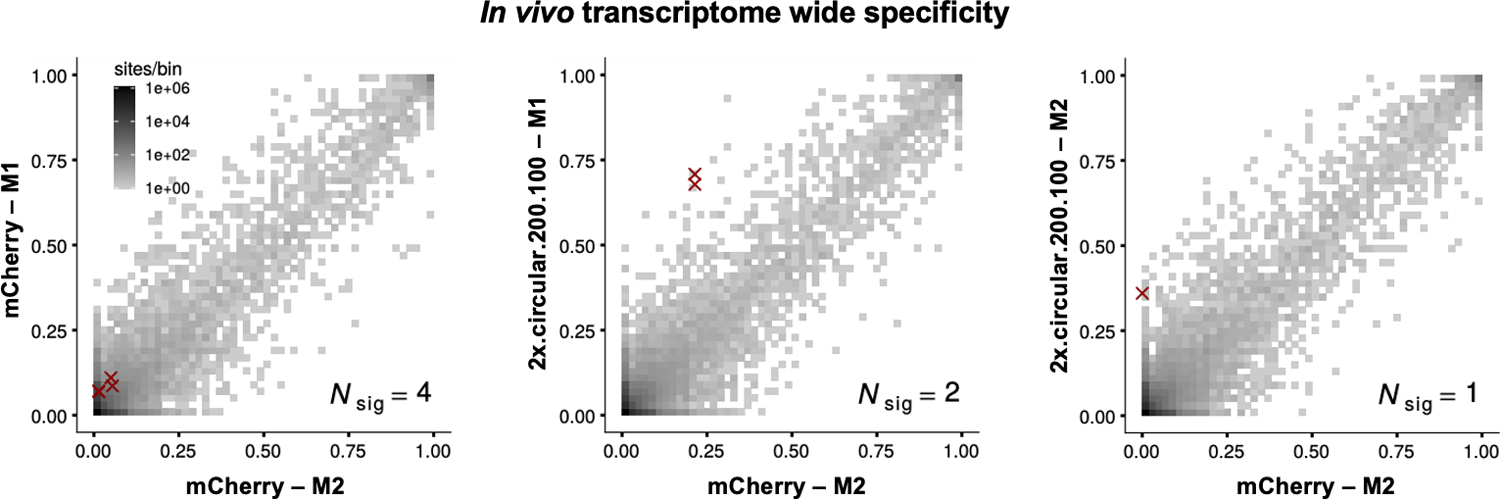
Comment in
-
In vivo RNA base editing with circular RNAs.Nat Rev Genet. 2022 Apr;23(4):196-197. doi: 10.1038/s41576-022-00463-y. Nat Rev Genet. 2022. PMID: 35194183 No abstract available.
References
-
- Melcher T et al. A mammalian RNA editing enzyme. Nature 379, 460–464 (1996). - PubMed
-
- Bass BL & Weintraub H An unwinding activity that covalently modifies its double-stranded RNA substrate. Cell 55, 1089–1098 (1988). - PubMed
-
- Bass BL & Weintraub H A developmentally regulated activity that unwinds RNA duplexes. Cell 48, 607–613 (1987). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
