

Prickle isoform participation in distinct polarization events in the Drosophila eye
- PMID: 35148314
- PMCID: PMC8836327
- DOI: 10.1371/journal.pone.0262328
Prickle isoform participation in distinct polarization events in the Drosophila eye
Abstract
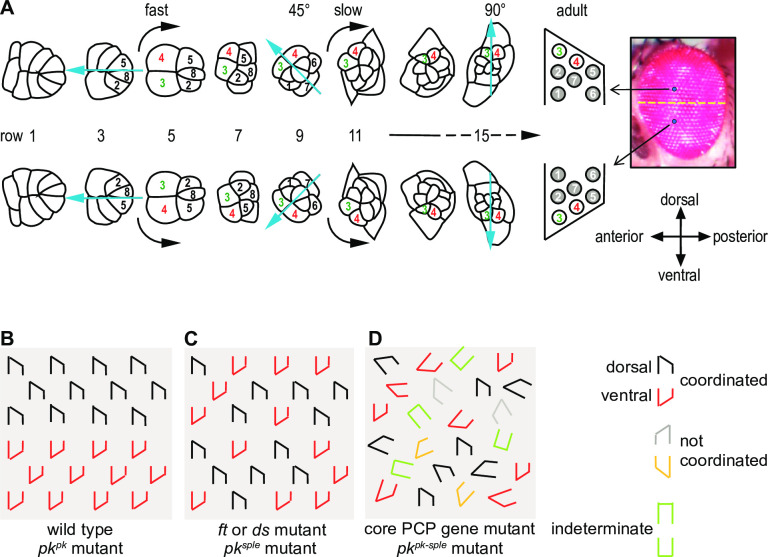
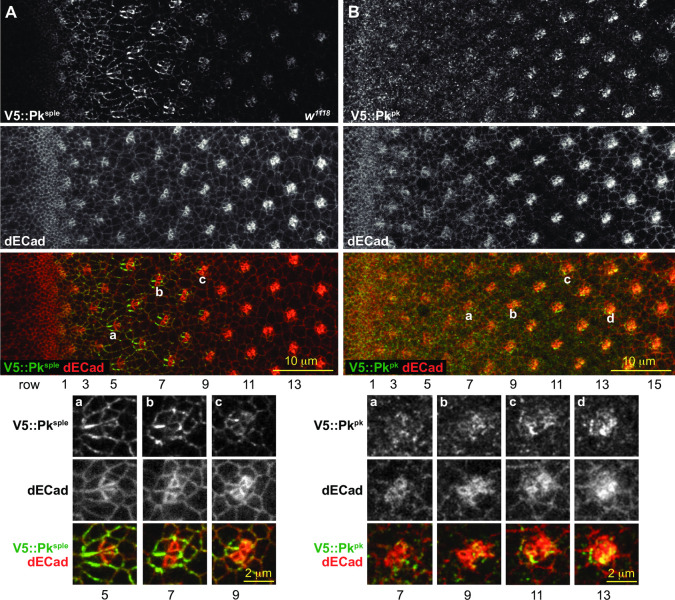
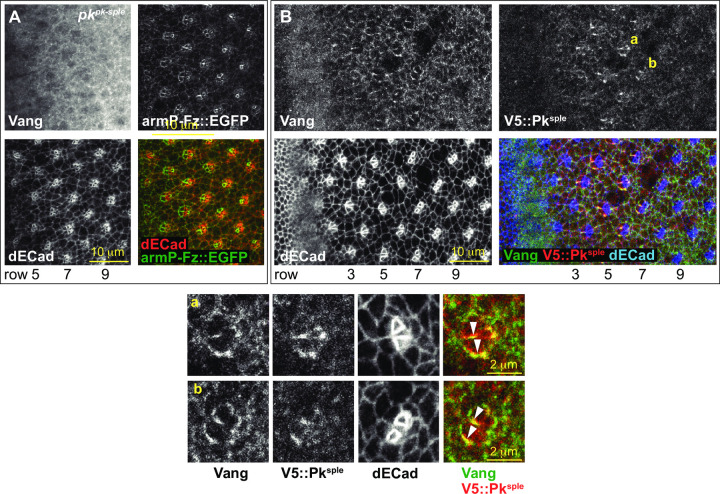
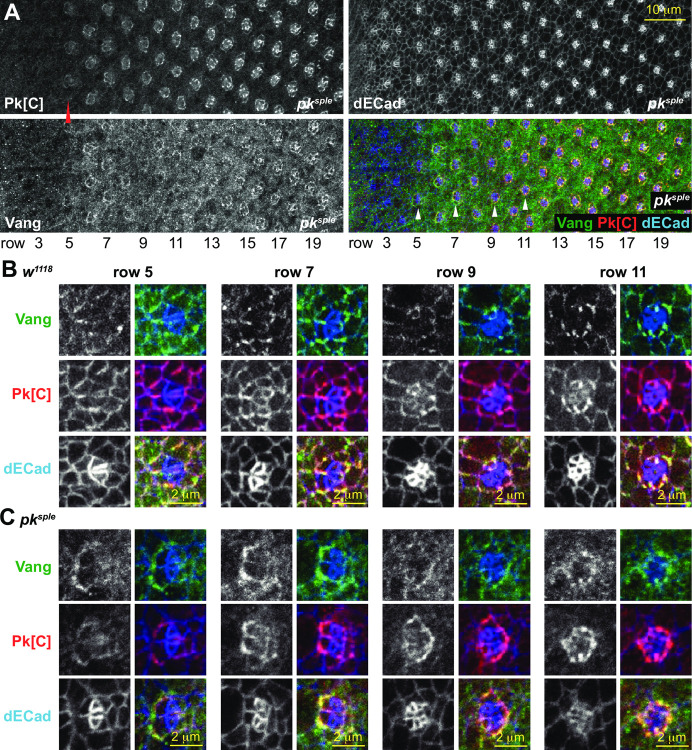
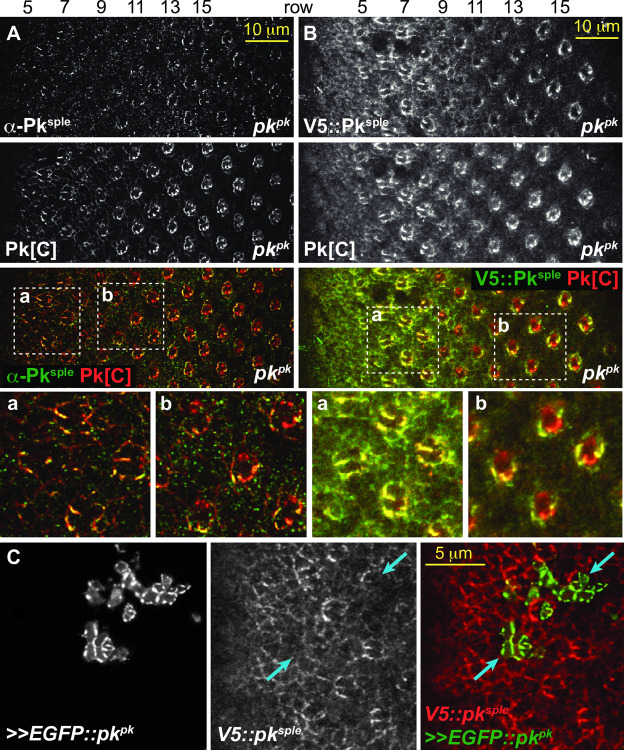
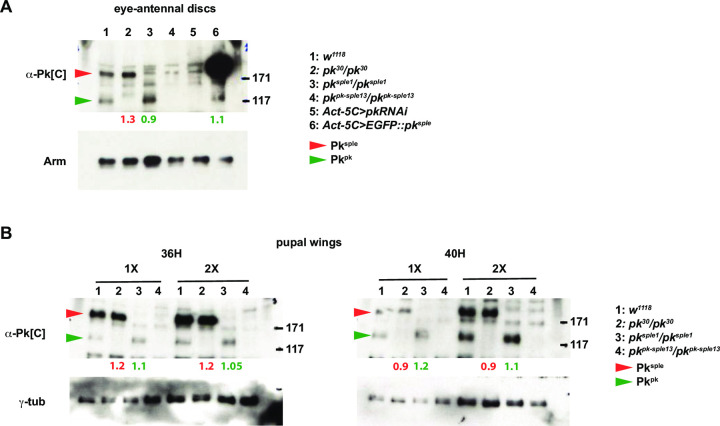
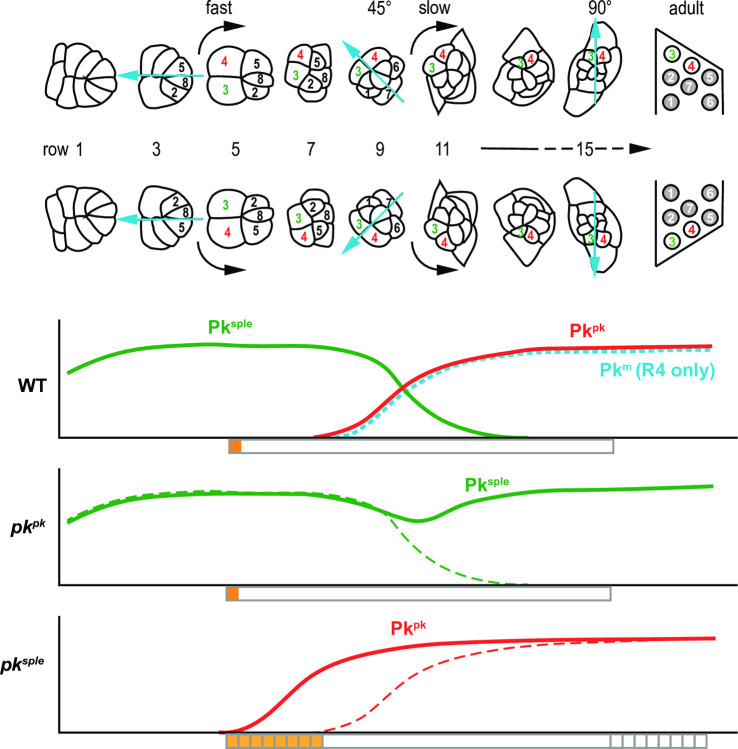
Planar cell polarity (PCP) signaling regulates several polarization events during development of ommatidia in the Drosophila eye, including directing chirality by polarizing a cell fate choice and determining the direction and extent of ommatidial rotation. The pksple isoform of the PCP protein Prickle is known to participate in the R3/R4 cell fate decision, but the control of other polarization events and the potential contributions of the three Pk isoforms have not been clarified. Here, by characterizing expression and subcellular localization of individual isoforms together with re-analyzing isoform specific phenotypes, we show that the R3/R4 fate decision, its coordination with rotation direction, and completion of rotation to a final ±90° rotation angle are separable polarization decisions with distinct Pk isoform requirements and contributions. Both pksple and pkpk can enforce robust R3/R4 fate decisions, but only pksple can correctly orient them along the dorsal-ventral axis. In contrast, pksple and pkpk can fully and interchangeably sustain coordination of rotation direction and rotation to completion. We propose that expression dynamics and competitive interactions determine isoform participation in these processes. We propose that the selective requirement for pksple to orient the R3/R4 decision and their interchangeability for coordination and completion of rotation reflects their previously described differential interaction with the Fat/Dachsous system which is known to be required for orientation of R3/R4 decisions but not for coordination or completion of rotation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Prickle is phosphorylated by Nemo and targeted for degradation to maintain Prickle/Spiny-legs isoform balance during planar cell polarity establishment.PLoS Genet. 2018 May 14;14(5):e1007391. doi: 10.1371/journal.pgen.1007391. eCollection 2018 May. PLoS Genet. 2018. PMID: 29758044 Free PMC article.
-
Combinatorial signaling by the Frizzled/PCP and Egfr pathways during planar cell polarity establishment in the Drosophila eye.Dev Biol. 2008 Apr 1;316(1):110-23. doi: 10.1016/j.ydbio.2008.01.016. Epub 2008 Jan 26. Dev Biol. 2008. PMID: 18291359 Free PMC article.
-
Prickle isoforms determine handedness of helical morphogenesis.Elife. 2020 Jan 14;9:e51456. doi: 10.7554/eLife.51456. Elife. 2020. PMID: 31934858 Free PMC article.
-
How do the Fat-Dachsous and core planar polarity pathways act together and independently to coordinate polarized cell behaviours?Open Biol. 2021 Feb;11(2):200356. doi: 10.1098/rsob.200356. Epub 2021 Feb 10. Open Biol. 2021. PMID: 33561385 Free PMC article. Review.
-
Planar cell polarity signaling in the Drosophila eye.Curr Top Dev Biol. 2010;93:189-227. doi: 10.1016/B978-0-12-385044-7.00007-2. Curr Top Dev Biol. 2010. PMID: 20959167 Free PMC article. Review.
Cited by
-
Distinct overlapping functions for Prickle1 and Prickle2 in the polarization of the airway epithelium.Front Cell Dev Biol. 2022 Sep 13;10:976182. doi: 10.3389/fcell.2022.976182. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36176272 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
