

Liquid-liquid phase separation drives cellular function and dysfunction in cancer
- PMID: 35149762
- PMCID: PMC10036213
- DOI: 10.1038/s41568-022-00444-7
Liquid-liquid phase separation drives cellular function and dysfunction in cancer
Abstract
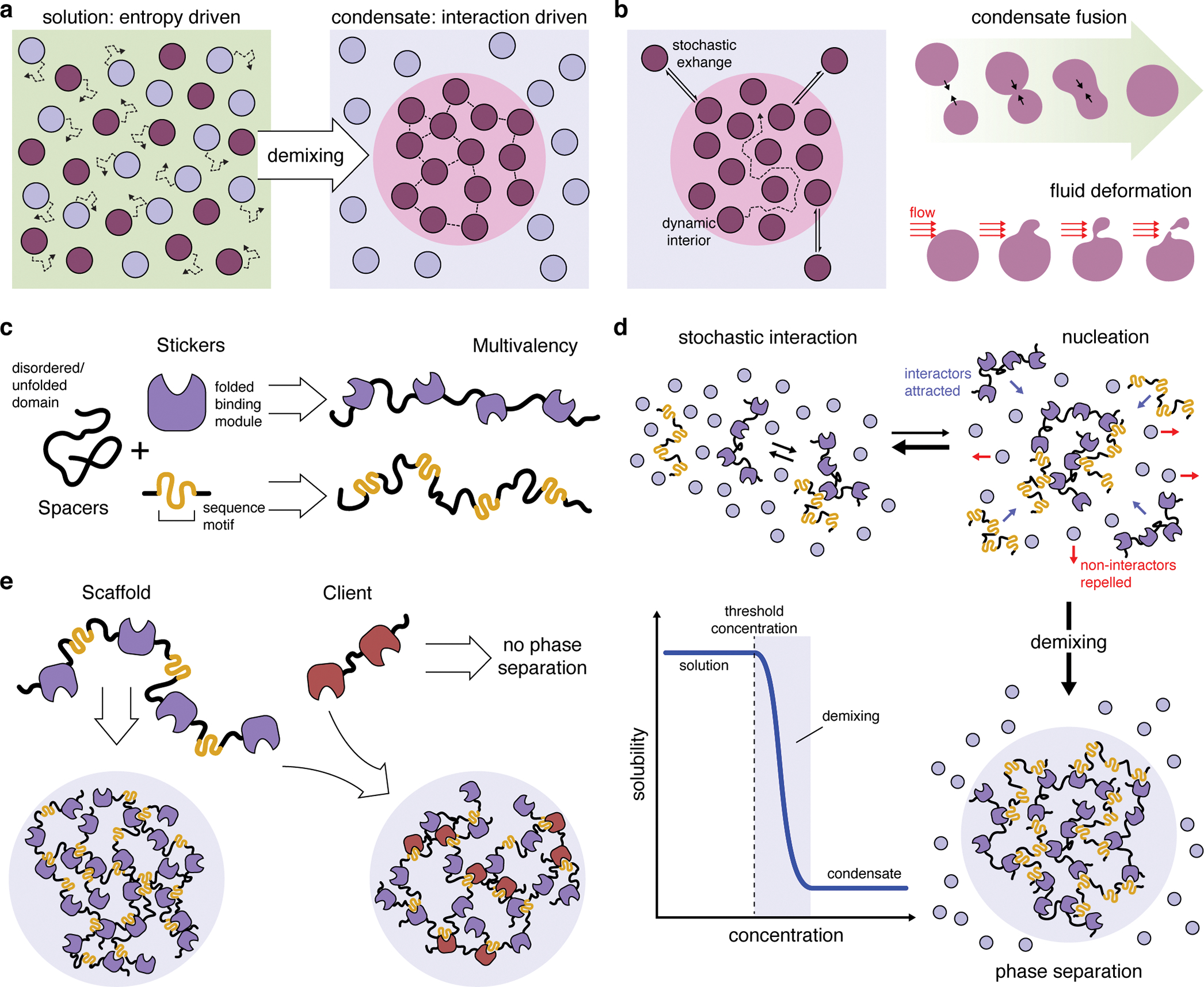
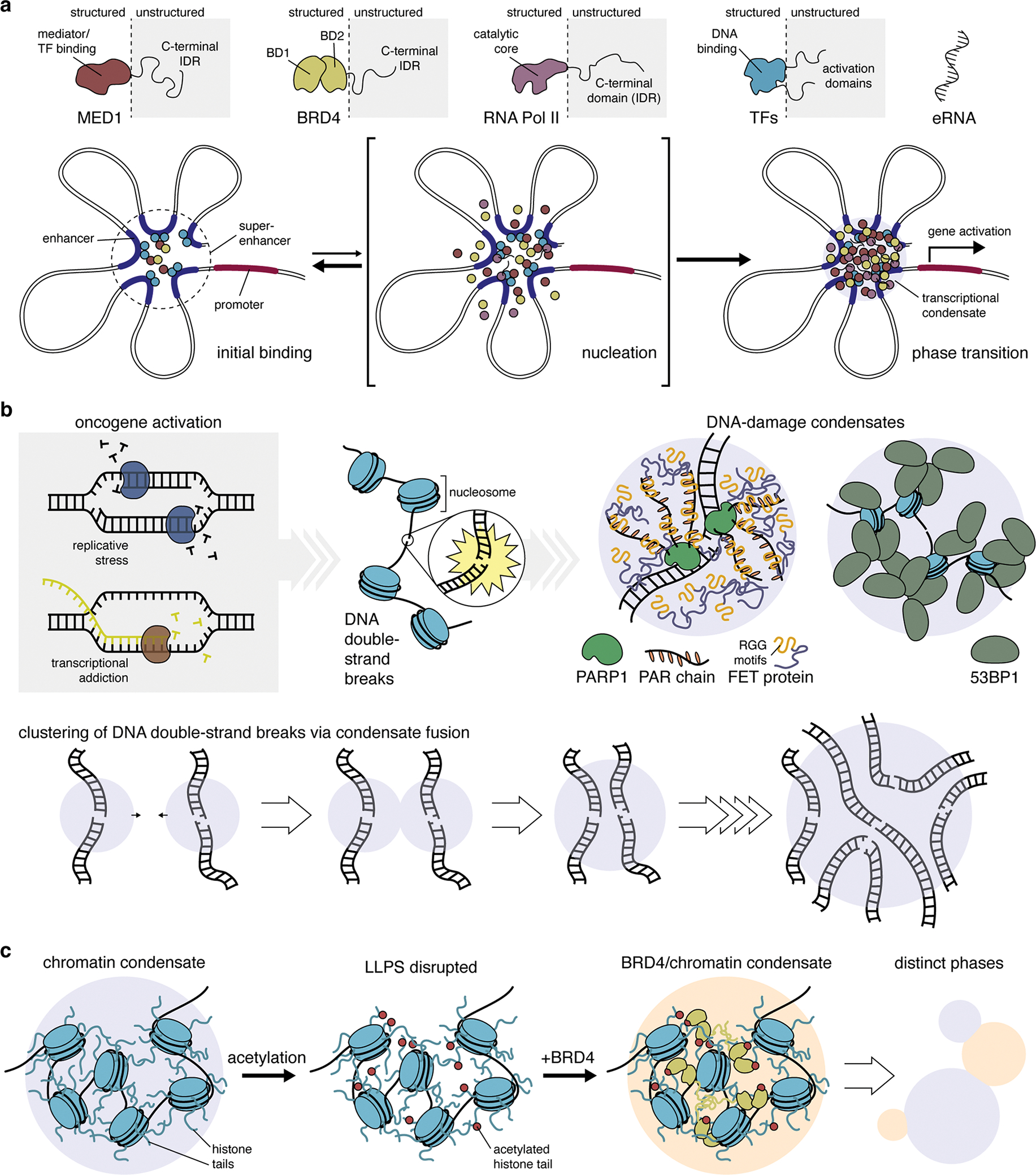
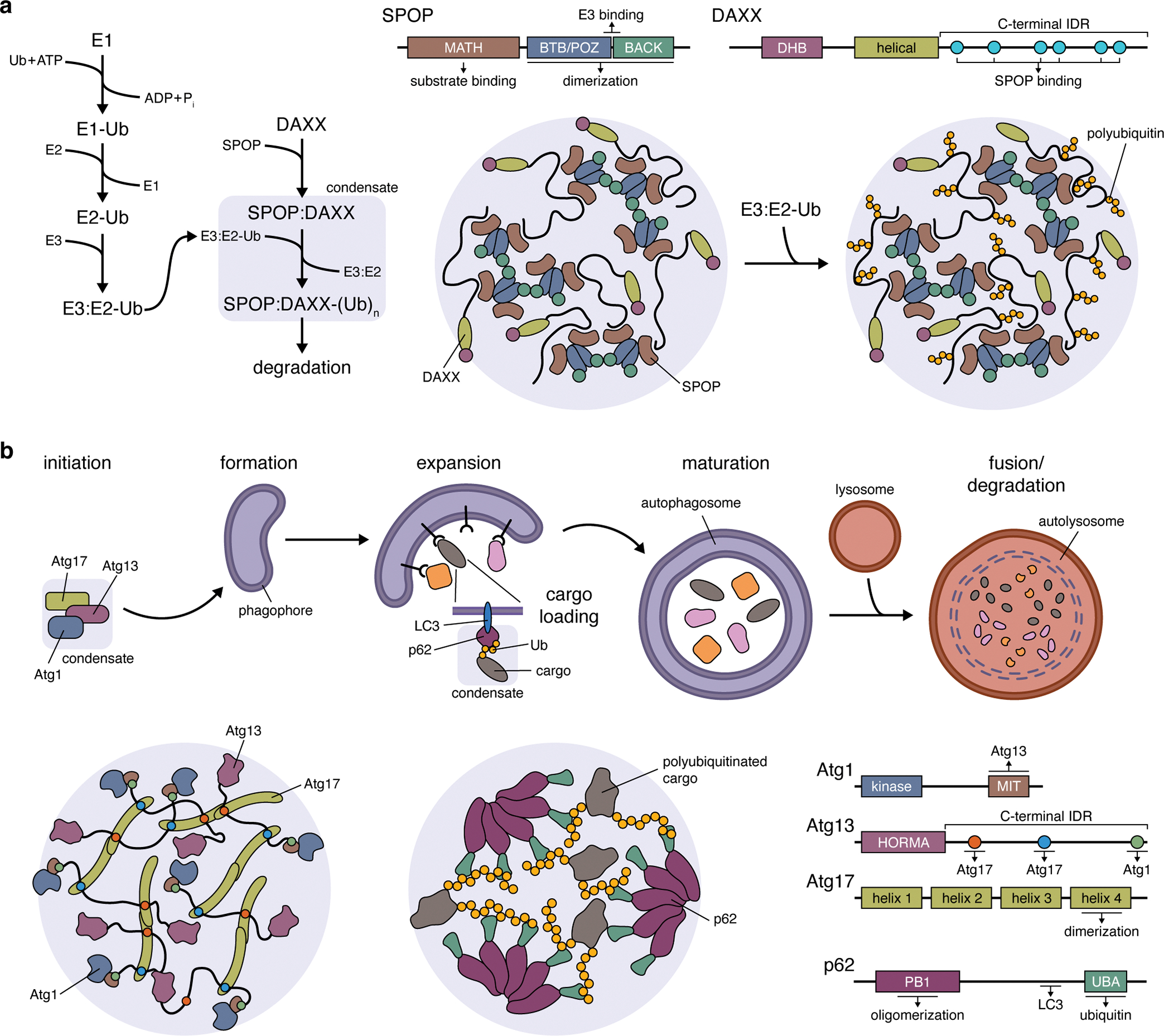
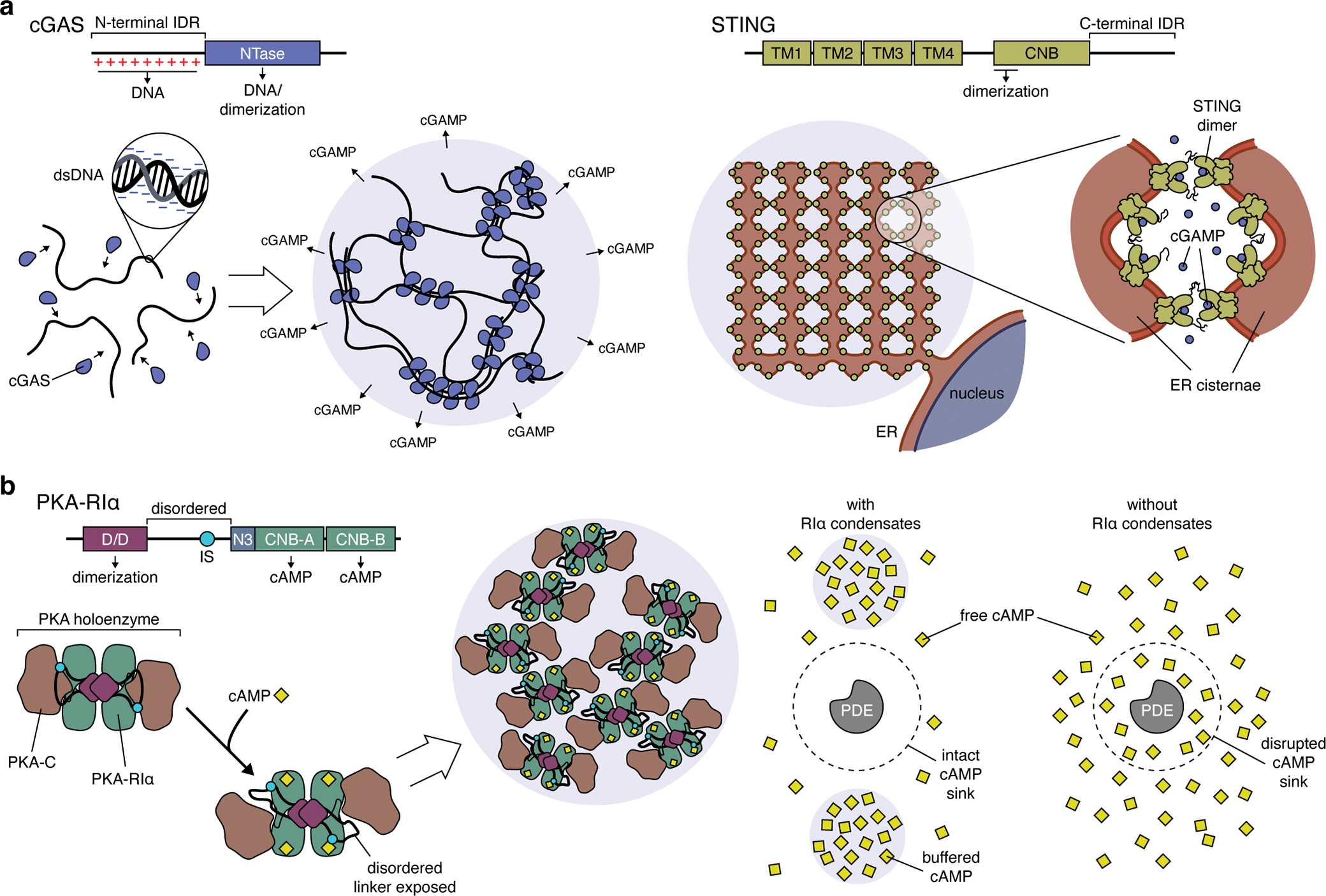
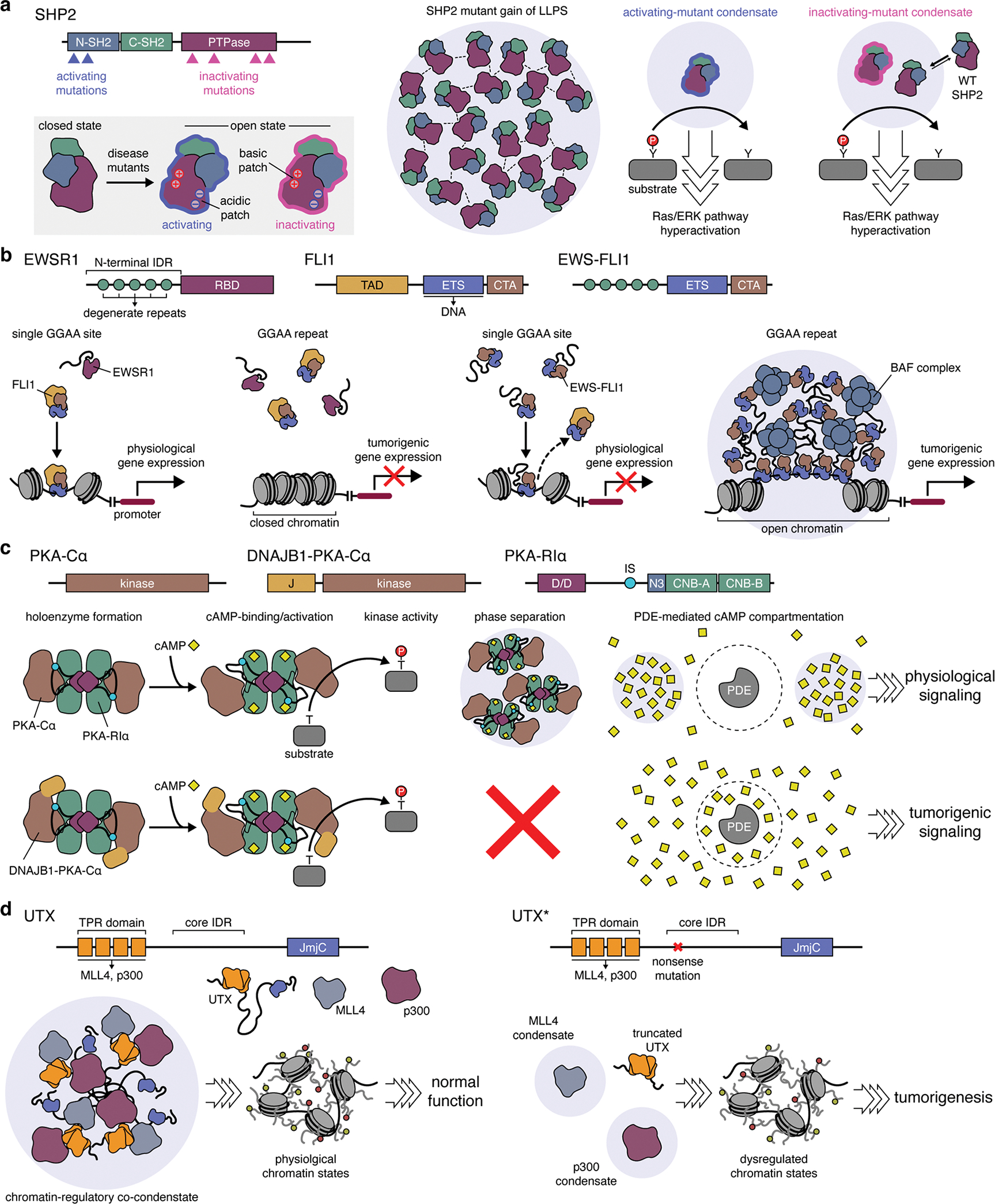
Cancer is a disease of uncontrollably reproducing cells. It is governed by biochemical pathways that have escaped the regulatory bounds of normal homeostatic balance. This balance is maintained through precise spatiotemporal regulation of these pathways. The formation of biomolecular condensates via liquid-liquid phase separation (LLPS) has recently emerged as a widespread mechanism underlying the spatiotemporal coordination of biological activities in cells. Biomolecular condensates are widely observed to directly regulate key cellular processes involved in cancer cell pathology, and the dysregulation of LLPS is increasingly implicated as a previously hidden driver of oncogenic activity. In this Perspective, we discuss how LLPS shapes the biochemical landscape of cancer cells.
© 2022. Springer Nature Limited.
Conflict of interest statement
COMPETING INTERESTS
The authors declare they have no competing interests.
Figures
References
-
- Lengauer C, Kinzler KW & Vogelstein B Genetic instabilities in human cancers. Nature 396, 643–649 (1998). - PubMed
