

Purine-Induced IFN-γ Promotes Uric Acid Production by Upregulating Xanthine Oxidoreductase Expression
- PMID: 35154100
- PMCID: PMC8829549
- DOI: 10.3389/fimmu.2022.773001
Purine-Induced IFN-γ Promotes Uric Acid Production by Upregulating Xanthine Oxidoreductase Expression
Abstract
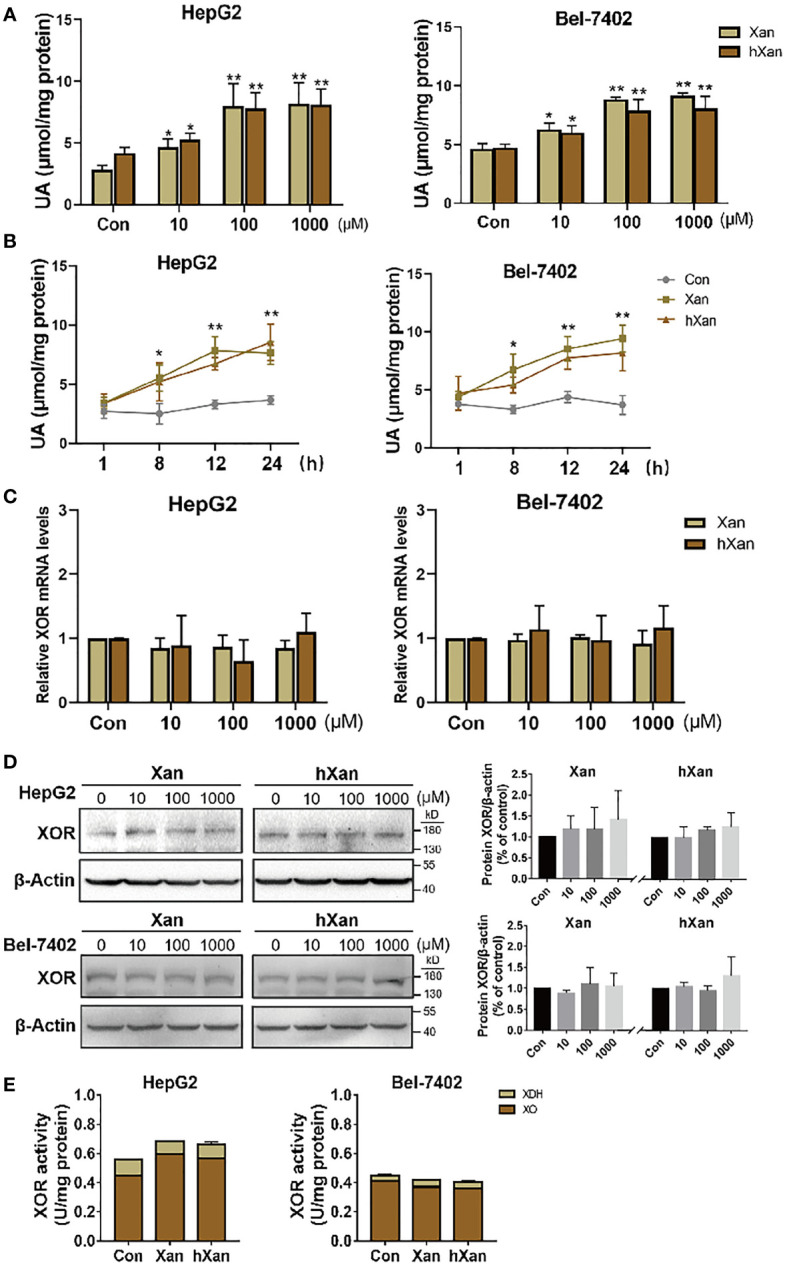
Objective: Limiting purine intake, inhibiting xanthine oxidoreductase (XOR) and inhibiting urate reabsorption in proximal tubule by uricosuric drugs, to reduce serum uric acid (UA) levels, are recognized treatments for gout. However, the mechanism of increased how XOR expression and activity in hyperuricemia and gout remains unclear. This study aims to explore whether exogenous purines are responsible for increased XOR expression and activity.
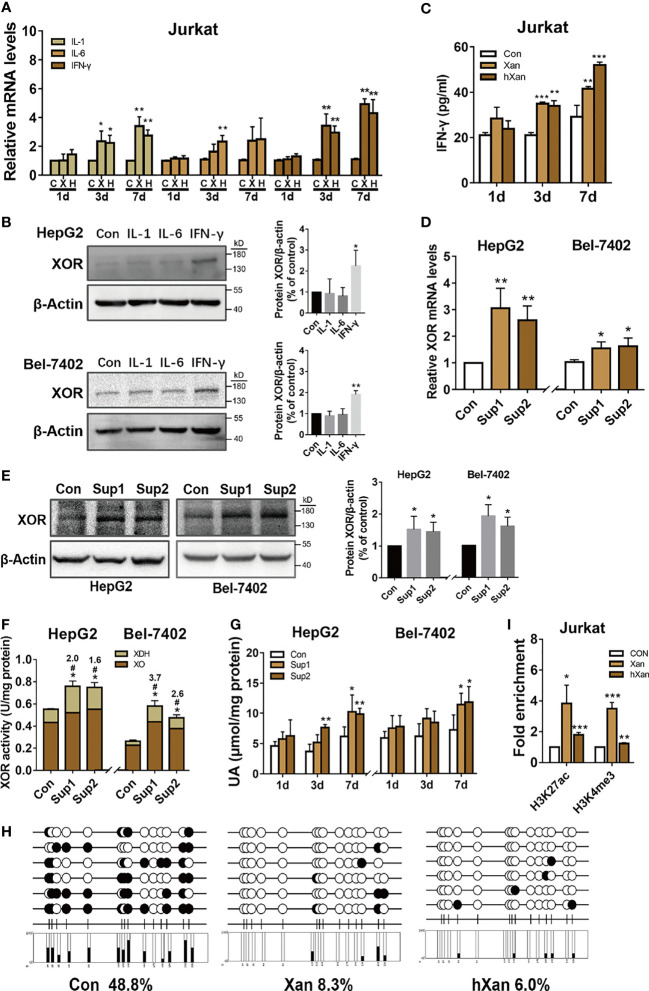
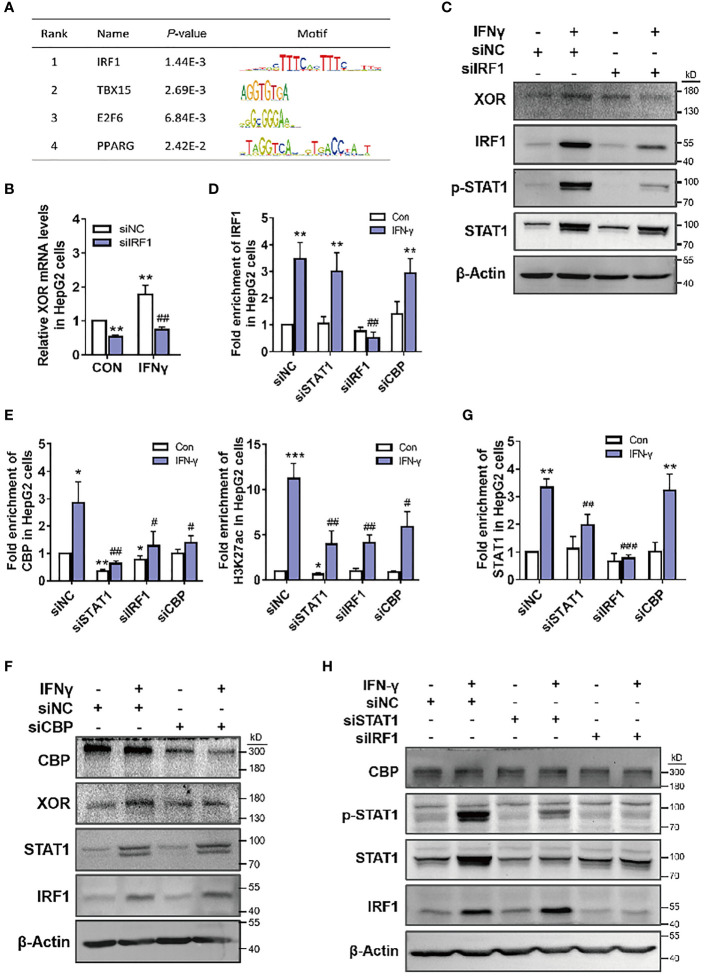
Methods: HepG2 and Bel-7402 human hepatoma cells were stimulated with exogenous purine, or were exposed to conditioned growth medium of purine-stimulated Jurkat cells, followed by measurement of XOR expression and UA production to determine the effect of lymphocyte-secreted cytokines on XOR expression in hepatocytes. The expression of STAT1, IRF1 and CBP and their binding on the XDH promoter were detected by western blotting and ChIP-qPCR. The level of DNA methylation was determined by bisulfite sequencing PCR. Blood samples from 117 hyperuricemia patients and 119 healthy individuals were collected to analyze the correlation between purine, UA and IFN-γ concentrations.
Results: Excess of purine was metabolized to UA in hepatocyte metabolism by XOR that was induced by IFN-γ secreted in the conditioned growth medium of Jurkat cells in response to exogenous purine, but it did not directly induce XOR expression. IFN-γ upregulated XOR expression due to the enhanced binding of STAT1 to IRF1 to further recruit CBP to the XDH promoter. Clinical data showed positive correlation of serum IFN-γ with both purine and UA, and associated risk of hyperuricemia.
Conclusion: Purine not only acts as a metabolic substrate of XOR for UA production, but it induces inflammation through IFN-γ secretion that stimulates UA production through elevation of XOR expression.
Keywords: IFN-γ; IRF1; STAT1; hyperuricemia; purine; xanthine oxidoreductase.
Copyright © 2022 Wang, Xie, Song, Wang, Li, Lin, Su, Liang and Huang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous