

Metnase and EEPD1: DNA Repair Functions and Potential Targets in Cancer Therapy
- PMID: 35155245
- PMCID: PMC8831698
- DOI: 10.3389/fonc.2022.808757
Metnase and EEPD1: DNA Repair Functions and Potential Targets in Cancer Therapy
Abstract
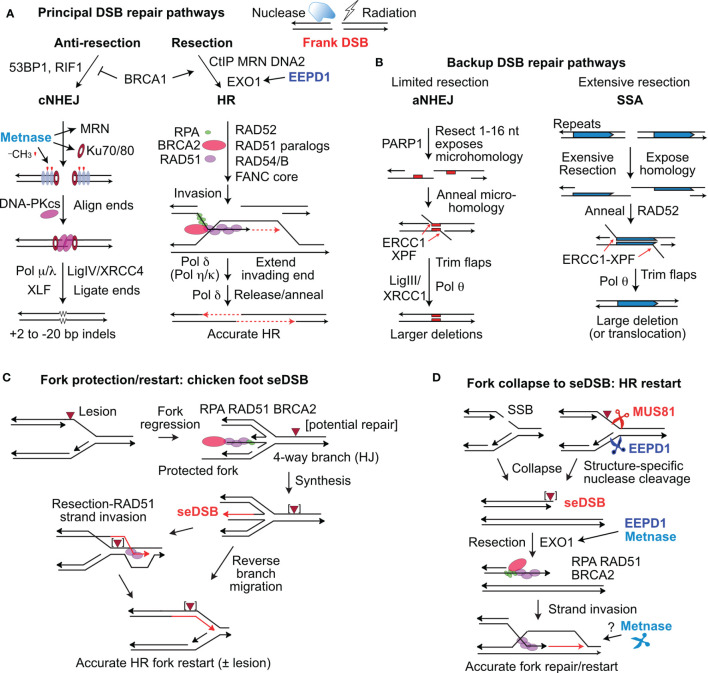
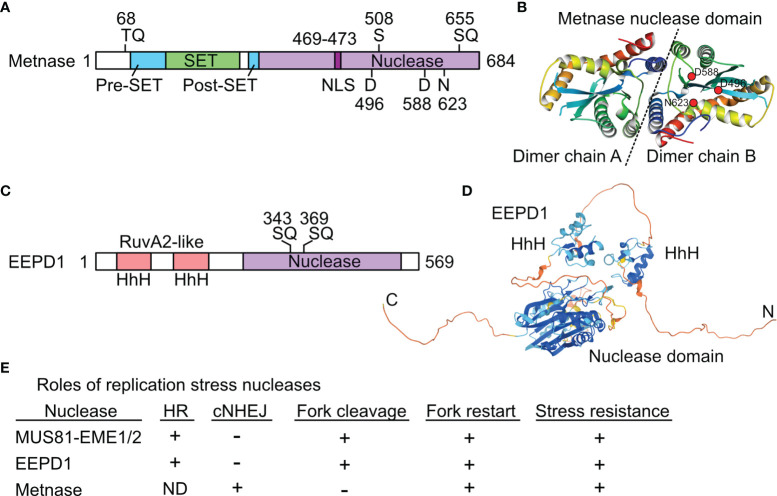
Cells respond to DNA damage by activating signaling and DNA repair systems, described as the DNA damage response (DDR). Clarifying DDR pathways and their dysregulation in cancer are important for understanding cancer etiology, how cancer cells exploit the DDR to survive endogenous and treatment-related stress, and to identify DDR targets as therapeutic targets. Cancer is often treated with genotoxic chemicals and/or ionizing radiation. These agents are cytotoxic because they induce DNA double-strand breaks (DSBs) directly, or indirectly by inducing replication stress which causes replication fork collapse to DSBs. EEPD1 and Metnase are structure-specific nucleases, and Metnase is also a protein methyl transferase that methylates histone H3 and itself. EEPD1 and Metnase promote repair of frank, two-ended DSBs, and both promote the timely and accurate restart of replication forks that have collapsed to single-ended DSBs. In addition to its roles in HR, Metnase also promotes DSB repair by classical non-homologous recombination, and chromosome decatenation mediated by TopoIIα. Although mutations in Metnase and EEPD1 are not common in cancer, both proteins are frequently overexpressed, which may help tumor cells manage oncogenic stress or confer resistance to therapeutics. Here we focus on Metnase and EEPD1 DNA repair pathways, and discuss opportunities for targeting these pathways to enhance cancer therapy.
Keywords: DNA damage; DNA double-strand breaks; DNA repair; chromosome decatenation; genome instability; homologous recombination; non-homologous end-joining.
Copyright © 2022 Nickoloff, Sharma, Taylor, Allen, Lee and Hromas.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The reviewer EM declared a shared affiliation, with one of the authors, S-HL, to the handling editor at the time of the review.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources