

Evolution of CDK1 Paralog Specializations in a Lineage With Fast Developing Planktonic Embryos
- PMID: 35155443
- PMCID: PMC8832800
- DOI: 10.3389/fcell.2021.770939
Evolution of CDK1 Paralog Specializations in a Lineage With Fast Developing Planktonic Embryos
Abstract
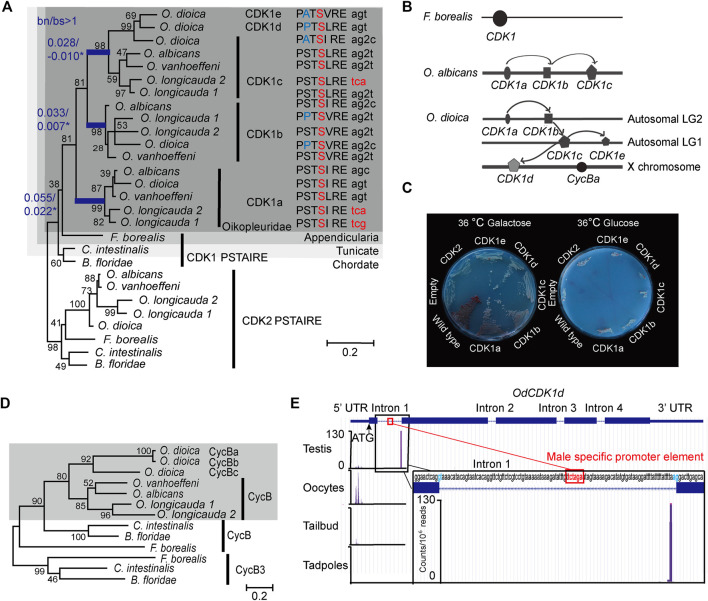
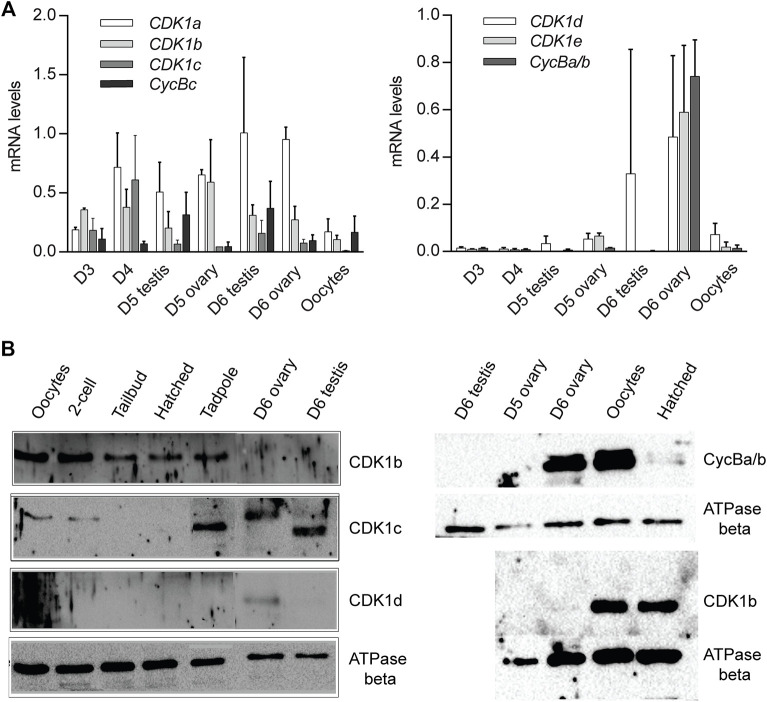
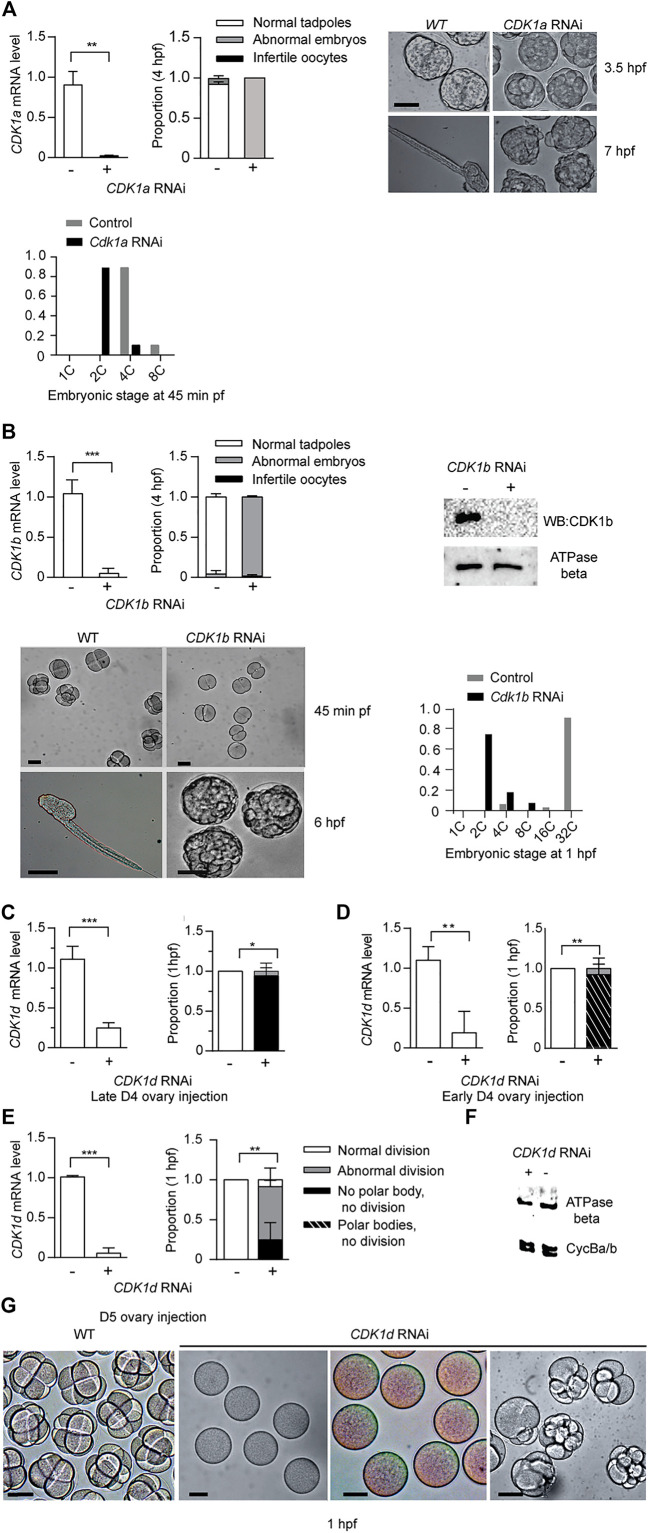
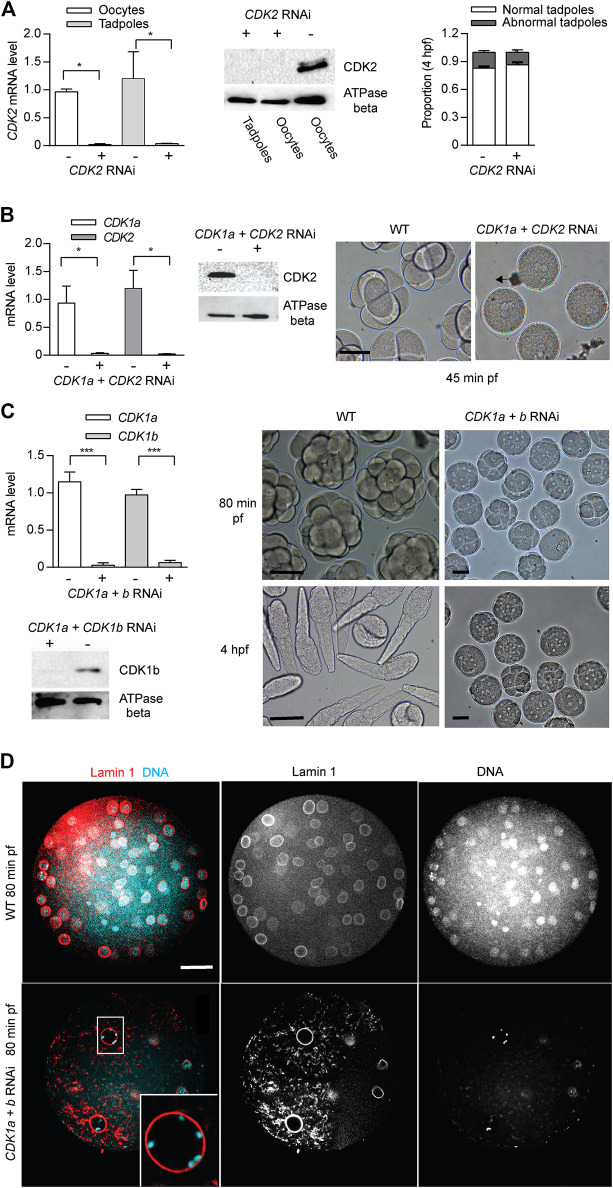
The active site of the essential CDK1 kinase is generated by core structural elements, among which the PSTAIRE motif in the critical αC-helix, is universally conserved in the single CDK1 ortholog of all metazoans. We report serial CDK1 duplications in the chordate, Oikopleura. Paralog diversifications in the PSTAIRE, activation loop substrate binding platform, ATP entrance site, hinge region, and main Cyclin binding interface, have undergone positive selection to subdivide ancestral CDK1 functions along the S-M phase cell cycle axis. Apparent coevolution of an exclusive CDK1d:Cyclin Ba/b pairing is required for oogenic meiosis and early embryogenesis, a period during which, unusually, CDK1d, rather than Cyclin Ba/b levels, oscillate, to drive very rapid cell cycles. Strikingly, the modified PSTAIRE of odCDK1d shows convergence over great evolutionary distance with plant CDKB, and in both cases, these variants exhibit increased specialization to M-phase.
Keywords: PSTAIRE helix; coenocyst; cyclin dependent kinase; determinant of specificity; embryogenesis; gene duplication; nuclear envelope breakdown; tunicate.
Copyright © 2022 Ma, Øvrebø and Thompson.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Specialization of CDK1 and cyclin B paralog functions in a coenocystic mode of oogenic meiosis.Cell Cycle. 2018;17(12):1425-1444. doi: 10.1080/15384101.2018.1486167. Epub 2018 Jul 23. Cell Cycle. 2018. PMID: 29969934 Free PMC article.
-
Expansion of cyclin D and CDK1 paralogs in Oikopleura dioica, a chordate employing diverse cell cycle variants.Mol Biol Evol. 2012 Feb;29(2):487-502. doi: 10.1093/molbev/msr136. Epub 2011 Jul 6. Mol Biol Evol. 2012. PMID: 21734012
-
Functional specialization of chordate CDK1 paralogs during oogenic meiosis.Cell Cycle. 2015;14(6):880-93. doi: 10.1080/15384101.2015.1006000. Cell Cycle. 2015. PMID: 25714331 Free PMC article.
-
Cyclin-dependent protein kinase inhibitors including palbociclib as anticancer drugs.Pharmacol Res. 2016 May;107:249-275. doi: 10.1016/j.phrs.2016.03.012. Epub 2016 Mar 16. Pharmacol Res. 2016. PMID: 26995305 Review.
-
Cyclin B1 and CDK1: nuclear localization and upstream regulators.Prog Cell Cycle Res. 2003;5:335-47. Prog Cell Cycle Res. 2003. PMID: 14593728 Review.
Cited by
-
Functional specialization of Aurora kinase homologs during oogenic meiosis in the tunicate Oikopleura dioica.Front Cell Dev Biol. 2023 Dec 7;11:1323378. doi: 10.3389/fcell.2023.1323378. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 38130951 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous