

A Novel In Situ Dendritic Cell Vaccine Triggered by Rose Bengal Enhances Adaptive Antitumour Immunity
- PMID: 35155685
- PMCID: PMC8824725
- DOI: 10.1155/2022/1178874
A Novel In Situ Dendritic Cell Vaccine Triggered by Rose Bengal Enhances Adaptive Antitumour Immunity
Retraction in
-
RETRACTION: A Novel In Situ Dendritic Cell Vaccine Triggered by Rose Bengal Enhances Adaptive Antitumour Immunity.J Immunol Res. 2025 Nov 13;2025:9834758. doi: 10.1155/jimr/9834758. eCollection 2025. J Immunol Res. 2025. PMID: 41245396 Free PMC article.
Abstract
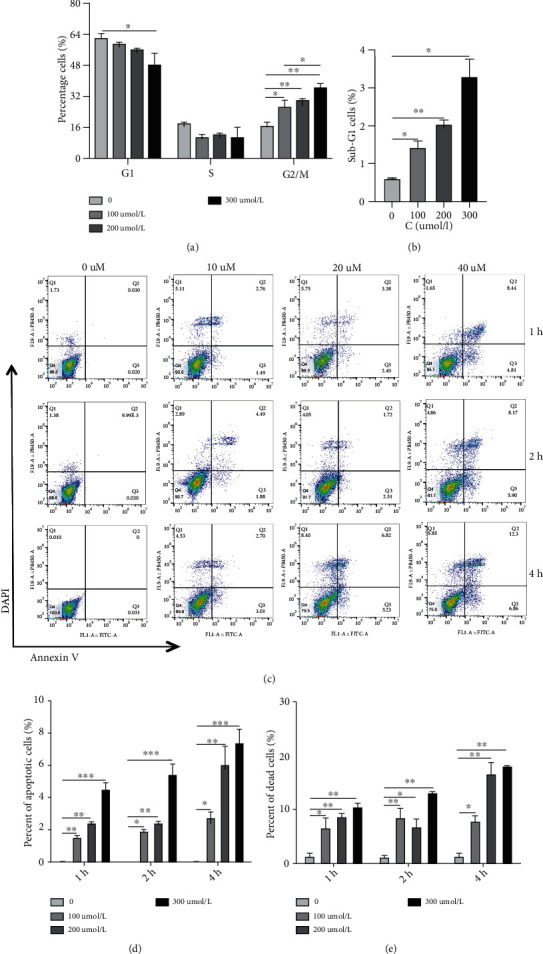
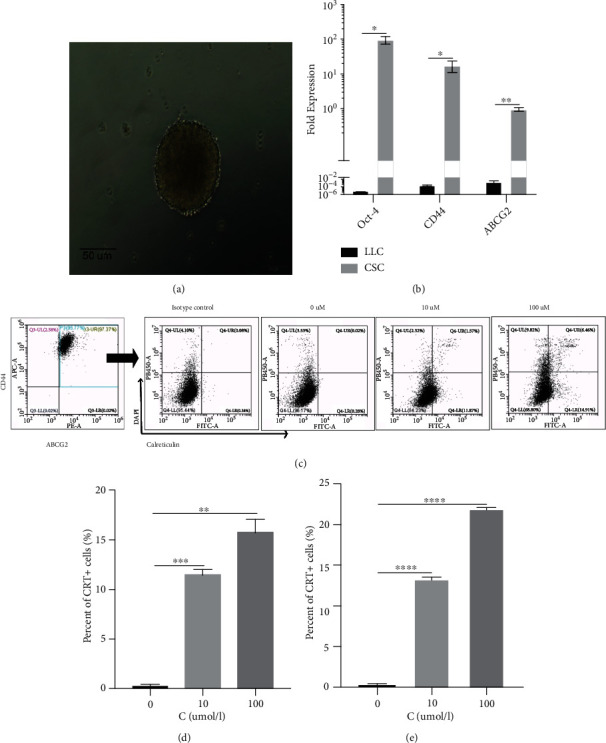
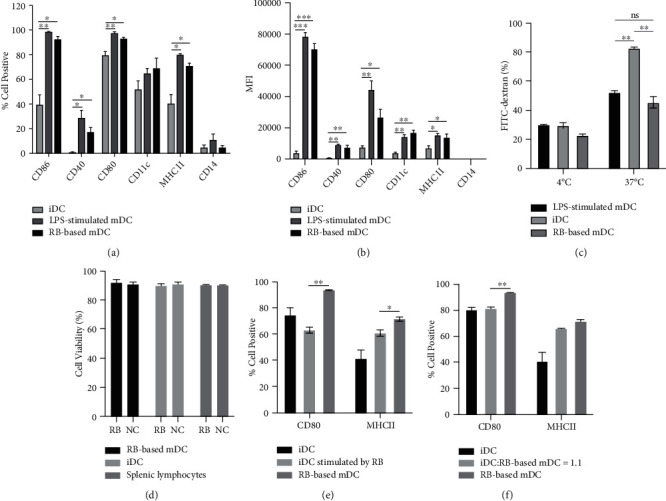
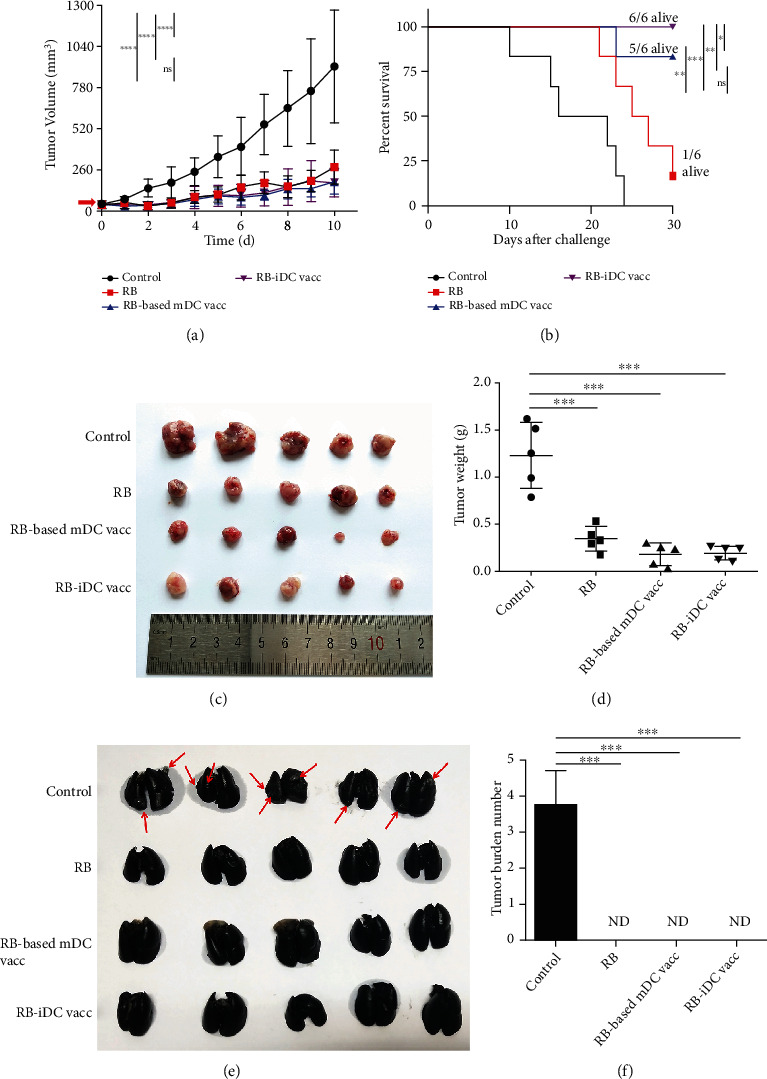
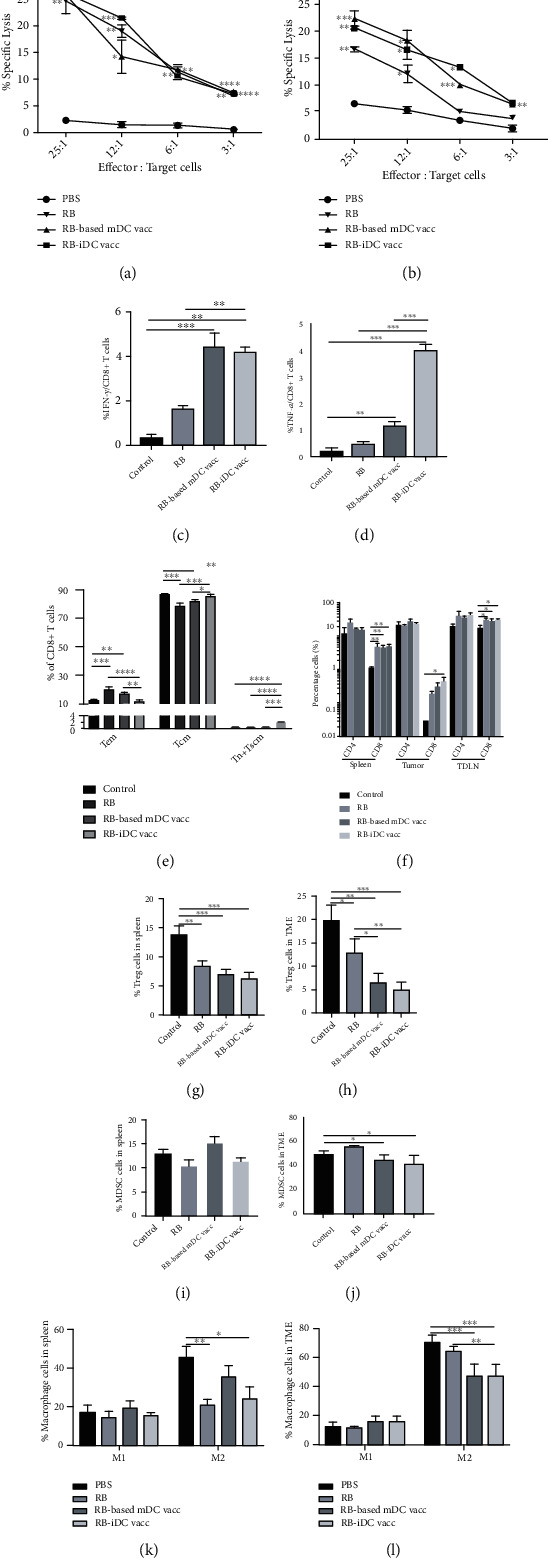
Dendritic cell- (DC-) based vaccination has emerged as a promising antitumour immunotherapy. However, overcoming immune tolerance and immunosuppression in the tumour microenvironment (TME) is still a great challenge. Recent studies have shown that Rose Bengal (RB) can effectively induce immunogenic cell death (ICD) in cancer cells, presenting whole tumour antigens for DC processing and presentation. However, the synergistic antitumour effect of combining intralesional RB with immature DCs (RB-iDCs) remains unclear. In the present study, we investigated whether RB-iDCs have superior antitumour effects compared with either single agent and evaluated the immunological mechanism of RB-iDCs in a murine lung cancer model. The results showed that intralesional RB-iDCs suppressed subcutaneous tumour growth and lung metastasis, which resulted in 100% mouse survival and significantly increased TNF-α production by CD8+ T cells. These effects were closely related to the induction of the expression of distinct ICD hallmarks by RB in both bulk cancer cells and cancer stem cells (CSCs), especially calreticulin (CRT), thus enhancing immune effector cell (i.e., CD4+, CD8+, and memory T cells) infiltration and attenuating the accumulation of immunosuppressive cells (i.e., Tregs, macrophages, and myeloid-derived suppressor cells (MDSCs)) in the TME. This study reveals that the RB-iDC vaccine can synergistically destroy the primary tumour, inhibit distant metastasis, and prevent tumour relapse in a lung cancer mouse model, which provides important preclinical data for the development of a novel combinatorial immunotherapy.
Copyright © 2022 Lanlin Zhang et al.
Conflict of interest statement
The authors declare no potential conflicts of interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
