

Sexual dimorphism: increased sterol excretion leads to hypocholesterolaemia in female hyperbilirubinaemic Gunn rats
- PMID: 35156712
- PMCID: PMC9310728
- DOI: 10.1113/JP282395
Sexual dimorphism: increased sterol excretion leads to hypocholesterolaemia in female hyperbilirubinaemic Gunn rats
Abstract
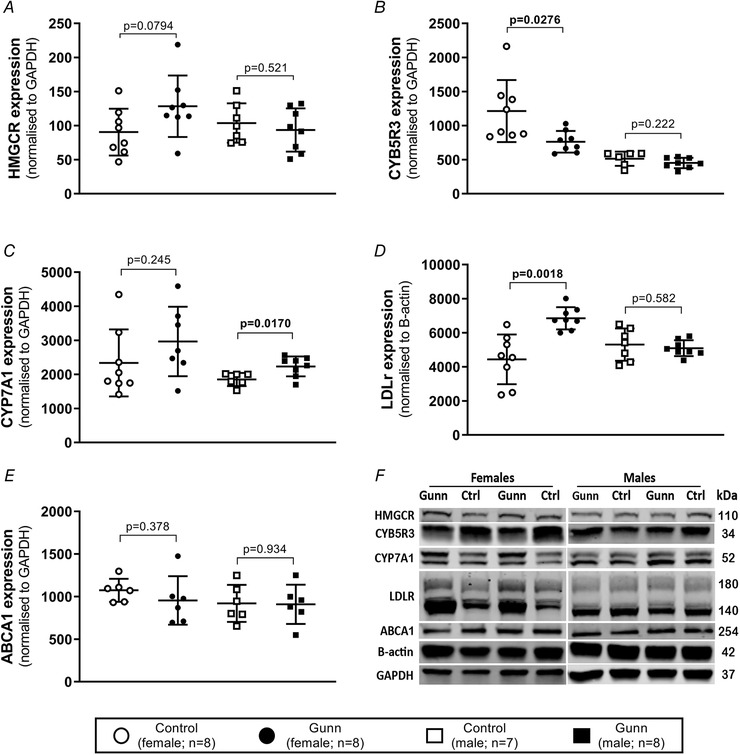
Circulating bilirubin is associated with reduced serum cholesterol concentrations in humans and in hyperbilirubinaemic Gunn rats. However, mechanisms contributing to hypocholesterolaemia remain unknown. Therefore, this study aimed to investigate cholesterol synthesis, transport and excretion in mutant Gunn rats. Adult Gunn and control rats were assessed for daily faecal sterol excretion using metabolic cages, and water was supplemented with [1-13 C]-acetate to determine cholesterol synthesis. Bile was collected to measure biliary lipid secretion. Serum and liver were collected for biochemical analysis and for gene/protein expression using RT-qPCR and western blot, respectively. Additionally, serum was collected and analysed from juvenile rats. A significant interaction of sex, age and phenotype on circulating lipids was found with adult female Gunn rats reporting significantly lower cholesterol and phospholipids. Female Gunn rats also demonstrated elevated cholesterol synthesis, greater biliary lipid secretion and increased total faecal cholesterol and bile acid excretion. Furthermore, they possessed increased hepatic low-density lipoprotein (LDL) receptor and SREBP2 expression. In contrast, there were no changes to sterol metabolism in adult male Gunn rats. This is the first study to demonstrate elevated faecal sterol excretion in female hyperbilirubinaemic Gunn rats. Increased sterol excretion creates a negative intestinal sterol balance that is compensated for by increased cholesterol synthesis and LDL receptor expression. Therefore, reduced circulating cholesterol is potentially caused by increased hepatic uptake via the LDL receptor. Future studies are required to further evaluate the sexual dimorphism of this response and whether similar findings occur in females with benign unconjugated hyperbilirubinaemia (Gilbert's syndrome). KEY POINTS: Female adult hyperbilirubinaemic (Gunn) rats demonstrated lower circulating cholesterol, corroborating human studies that report a negative association between bilirubin and cholesterol concentrations. Furthermore, female Gunn rats had elevated sterol excretion creating a negative intestinal sterol balance that was compensated for by elevated cholesterol synthesis and increased hepatic low-density lipoprotein (LDL) receptor expression. Therefore, elevated LDL receptor expression potentially leads to reduced circulating cholesterol levels in female Gunn rats providing an explanation for the hypocholesterolaemia observed in humans with elevated bilirubin levels. This study also reports a novel interaction of sex with the hyperbilirubinaemic phenotype on sterol metabolism because changes were only reported in females and not in male Gunn rats. Future studies are required to further evaluate the sexual dimorphism of this response and whether similar findings occur in females with benign unconjugated hyperbilirubinaemia (Gilbert's syndrome).
Keywords: Gilbert's syndrome; LDL receptor; SREBP; UGT1A1; bile acid metabolism; cholesterol metabolism; lipid; unconjugated bilirubin.
© 2022 The Authors. The Journal of Physiology published by John Wiley & Sons Ltd on behalf of The Physiological Society.
Figures








Similar articles
-
Protection from age-related increase in lipid biomarkers and inflammation contributes to cardiovascular protection in Gilbert's syndrome.Clin Sci (Lond). 2013 Sep;125(5):257-64. doi: 10.1042/CS20120661. Clin Sci (Lond). 2013. PMID: 23566065
-
Intestinal excretion of unconjugated bilirubin in man and rats with inherited unconjugated hyperbilirubinemia.Pediatr Res. 1997 Aug;42(2):195-200. doi: 10.1203/00006450-199708000-00011. Pediatr Res. 1997. PMID: 9262222
-
Induction of fecal cholesterol excretion is not effective for the treatment of hyperbilirubinemia in Gunn rats.Pediatr Res. 2021 Feb;89(3):510-517. doi: 10.1038/s41390-020-0926-2. Epub 2020 May 1. Pediatr Res. 2021. PMID: 32357361
-
Bilirubin and beyond: a review of lipid status in Gilbert's syndrome and its relevance to cardiovascular disease protection.Prog Lipid Res. 2013 Apr;52(2):193-205. doi: 10.1016/j.plipres.2012.11.001. Epub 2012 Nov 29. Prog Lipid Res. 2013. PMID: 23201182 Review.
-
Gilbert's syndrome and drug metabolism.Clin Pharmacokinet. 1979 May-Jun;4(3):223-32. doi: 10.2165/00003088-197904030-00004. Clin Pharmacokinet. 1979. PMID: 383356 Review.
Cited by
-
Trans-ancestral rare variant association study with machine learning-based phenotyping for metabolic dysfunction-associated steatotic liver disease.Genome Biol. 2025 Mar 10;26(1):50. doi: 10.1186/s13059-025-03518-5. Genome Biol. 2025. PMID: 40065360 Free PMC article.
-
Bilirubin Metabolism Does Not Influence Serum Bile Acid Profiles According to LC-MS: A Human Case-Control Study.Int J Mol Sci. 2025 Mar 10;26(6):2475. doi: 10.3390/ijms26062475. Int J Mol Sci. 2025. PMID: 40141122 Free PMC article.
References
-
- de Boer JF, Kuipers F & Groen AK (2018). Cholesterol transport revisited: a new turbo mechanism to drive cholesterol excretion. Trends Endocrinol Metab 29, 123–133. - PubMed
-
- Bulmer AC, Bakrania B, Du Toit EF, Boon AC, Clark PJ, Powell LW, Wagner KH & Headrick JP (2018). Bilirubin acts as a multipotent guardian of cardiovascular integrity: more than just a radical idea. Am J Physiol ‐ Hear Circ Physiol 315, H429–H447. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical