

Distinct roles of the dystrophin-glycoprotein complex: α-dystrobrevin and α-syntrophin in the maintenance of the postsynaptic apparatus of the neuromuscular synapse
- PMID: 35157076
- PMCID: PMC9307313
- DOI: 10.1093/hmg/ddac041
Distinct roles of the dystrophin-glycoprotein complex: α-dystrobrevin and α-syntrophin in the maintenance of the postsynaptic apparatus of the neuromuscular synapse
Abstract
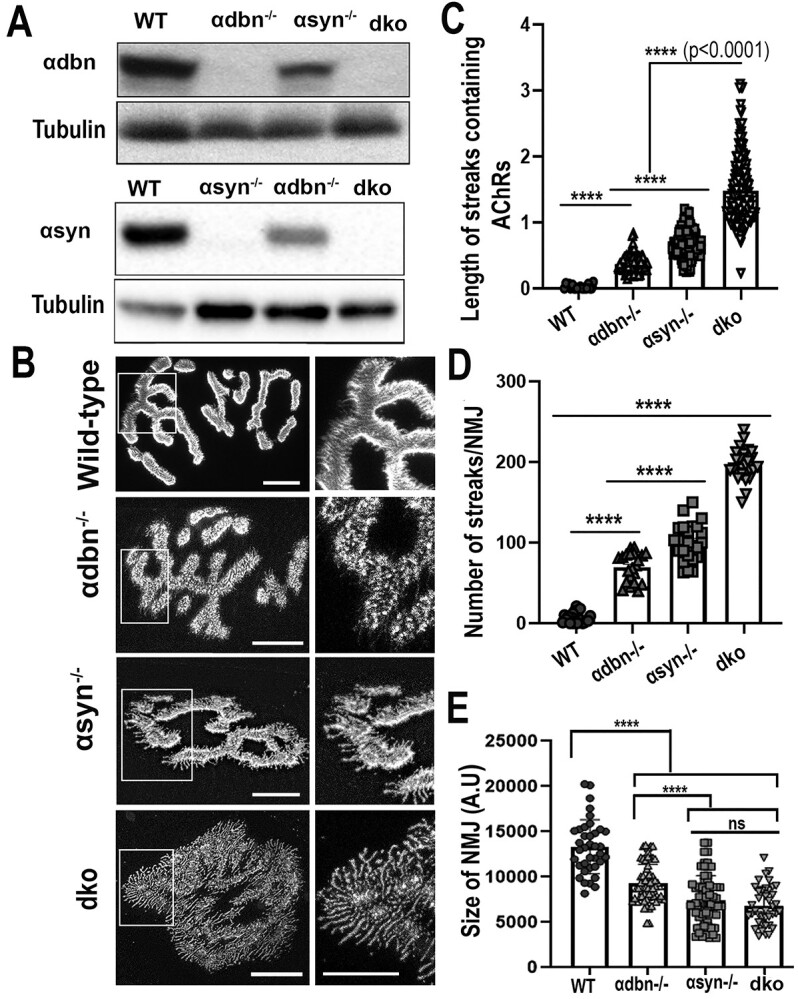
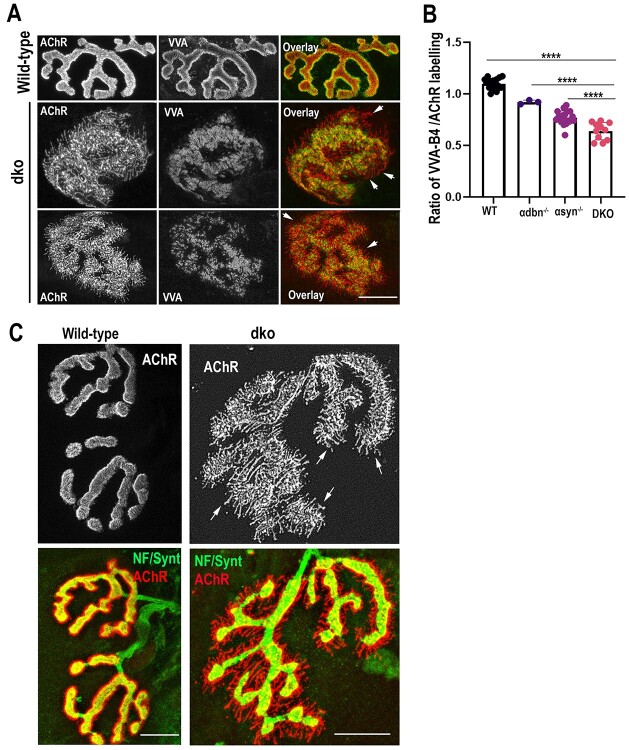
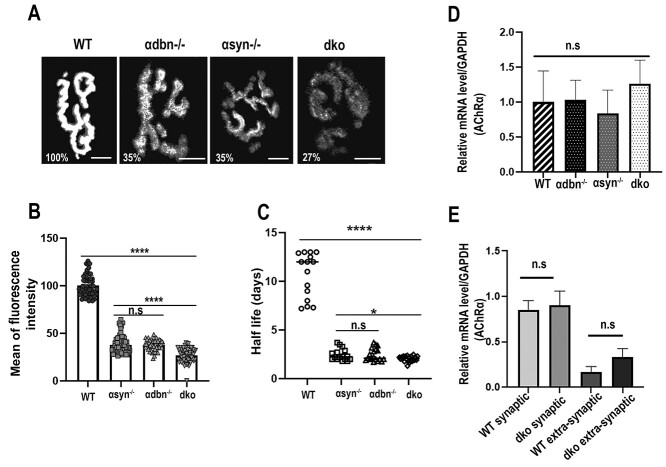
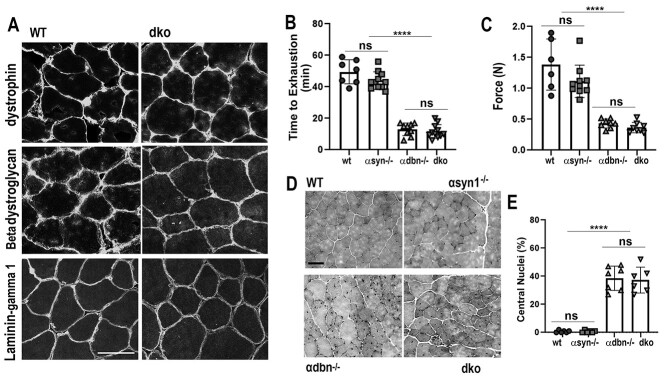
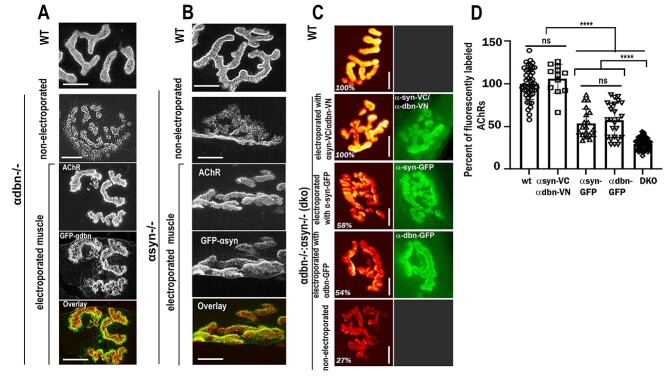
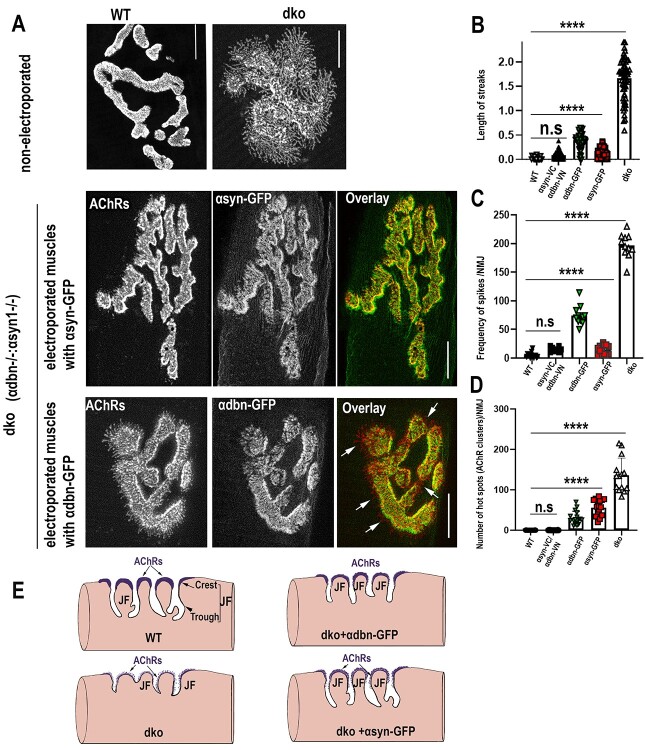
α-syntrophin (α-syn) and α-dystrobrevin (α-dbn), two components of the dystrophin-glycoprotein complex, are essential for the maturation and maintenance of the neuromuscular junction (NMJ) and mice deficient in either α-syn or α-dbn exhibit similar synaptic defects. However, the functional link between these two proteins and whether they exert distinct or redundant functions in the postsynaptic organization of the NMJ remain largely unknown. We generated and analyzed the synaptic phenotype of double heterozygote (α-dbn+/-, α-syn+/-), and double homozygote knockout (α-dbn-/-; α-syn-/-) mice and examined the ability of individual molecules to restore their defects in the synaptic phenotype. We showed that in double heterozygote mice, NMJs have normal synaptic phenotypes and no signs of muscular dystrophy. However, in double knockout mice (α-dbn-/-; α-syn-/-), the synaptic phenotype (the density, the turnover and the distribution of AChRs within synaptic branches) is more severely impaired than in single α-dbn-/- or α-syn-/- mutants. Furthermore, double mutant and single α-dbn-/- mutant mice showed more severe exercise-induced fatigue and more significant reductions in grip strength than single α-syn-/- mutant and wild-type. Finally, we showed that the overexpression of the transgene α-syn-GFP in muscles of double mutant restores primarily the abnormal extensions of membrane containing AChRs that extend beyond synaptic gutters and lack synaptic folds, whereas the overexpression of α-dbn essentially restores the abnormal dispersion of patchy AChR aggregates in the crests of synaptic folds. Altogether, these data suggest that α-syn and α-dbn act in parallel pathways and exert distinct functions on the postsynaptic structural organization of NMJs.
© The Author(s) 2022. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures

Similar articles
-
Nicotinic acetylcholine receptor stability at the NMJ deficient in α-syntrophin in vivo.J Neurosci. 2011 Oct 26;31(43):15586-96. doi: 10.1523/JNEUROSCI.4038-11.2011. J Neurosci. 2011. PMID: 22031904 Free PMC article.
-
Defects in neuromuscular junction structure in dystrophic muscle are corrected by expression of a NOS transgene in dystrophin-deficient muscles, but not in muscles lacking alpha- and beta1-syntrophins.Hum Mol Genet. 2004 Sep 1;13(17):1873-84. doi: 10.1093/hmg/ddh204. Epub 2004 Jul 6. Hum Mol Genet. 2004. PMID: 15238508
-
Spatial distribution and molecular dynamics of dystrophin glycoprotein components at the neuromuscular junction in vivo.J Cell Sci. 2017 May 15;130(10):1752-1759. doi: 10.1242/jcs.198358. Epub 2017 Mar 31. J Cell Sci. 2017. PMID: 28364093 Free PMC article.
-
The molecular cross talk of the dystrophin-glycoprotein complex.Ann N Y Acad Sci. 2018 Jan;1412(1):62-72. doi: 10.1111/nyas.13500. Epub 2017 Oct 25. Ann N Y Acad Sci. 2018. PMID: 29068540 Review.
-
The postsynaptic submembrane machinery at the neuromuscular junction: requirement for rapsyn and the utrophin/dystrophin-associated complex.J Neurocytol. 2003 Jun-Sep;32(5-8):709-26. doi: 10.1023/B:NEUR.0000020619.24681.2b. J Neurocytol. 2003. PMID: 15034263 Review.
Cited by
-
Plasticity and structural alterations of mitochondria and sarcoplasmic organelles in muscles of mice deficient in α-dystrobrevin, a component of the dystrophin-glycoprotein complex.Hum Mol Genet. 2024 Jun 21;33(13):1107-1119. doi: 10.1093/hmg/ddae039. Hum Mol Genet. 2024. PMID: 38507070 Free PMC article.
-
Morphological and functional alterations of neuromuscular synapses in a mouse model of ACTA1 congenital myopathy.Hum Mol Genet. 2024 Jan 20;33(3):233-244. doi: 10.1093/hmg/ddad183. Hum Mol Genet. 2024. PMID: 37883471 Free PMC article.
-
Regulation of miR-206 in denervated and dystrophic muscles, and its effect on acetylcholine receptor clustering.J Cell Sci. 2024 Dec 15;137(24):jcs262303. doi: 10.1242/jcs.262303. Epub 2024 Dec 13. J Cell Sci. 2024. PMID: 39575567
-
Nerve-independent formation of membrane infoldings at topologically complex postsynaptic apparatus by caveolin-3.Sci Adv. 2023 Jun 16;9(24):eadg0183. doi: 10.1126/sciadv.adg0183. Epub 2023 Jun 16. Sci Adv. 2023. PMID: 37327338 Free PMC article.
References
-
- Ervasti, J.M. (2007) Dystrophin, its interactions with other proteins, and implications for muscular dystrophy. Biochim. Biophys. Acta, 1772, 108–117. - PubMed
-
- Sunada, Y. and Campbell, K.P. (1995) Dystrophin-glycoprotein complex: molecular organization and critical roles in skeletal muscle. Curr. Opin. Neurol., 8, 379–384. - PubMed
-
- Durbeej, M. and Campbell, K.P. (2002) Muscular dystrophies involving the dystrophin-glycoprotein complex: an overview of current mouse models. Curr. Opin. Genet. Dev., 12, 349–361. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
