

Plant-microbe interactions in the apoplast: Communication at the plant cell wall
- PMID: 35157079
- PMCID: PMC9048882
- DOI: 10.1093/plcell/koac040
Plant-microbe interactions in the apoplast: Communication at the plant cell wall
Abstract
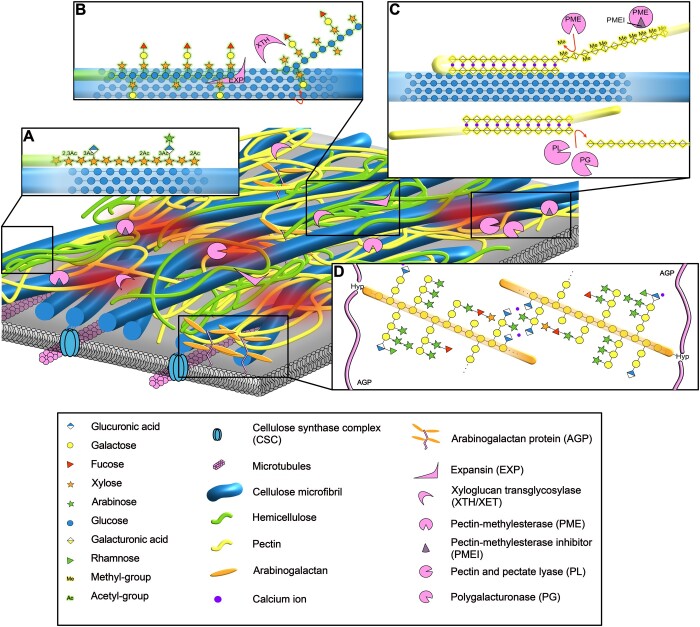
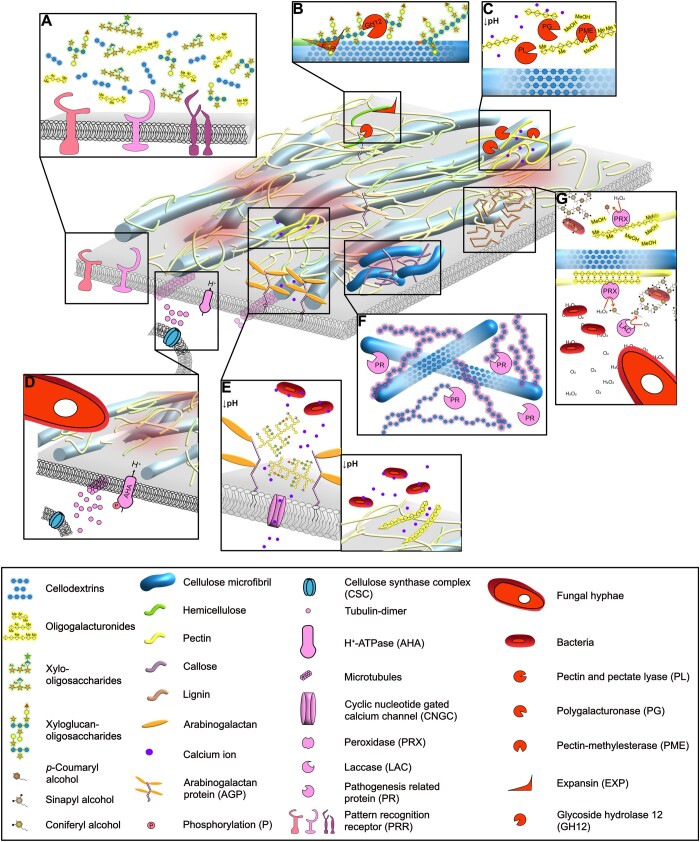
The apoplast is a continuous plant compartment that connects cells between tissues and organs and is one of the first sites of interaction between plants and microbes. The plant cell wall occupies most of the apoplast and is composed of polysaccharides and associated proteins and ions. This dynamic part of the cell constitutes an essential physical barrier and a source of nutrients for the microbe. At the same time, the plant cell wall serves important functions in the interkingdom detection, recognition, and response to other organisms. Thus, both plant and microbe modify the plant cell wall and its environment in versatile ways to benefit from the interaction. We discuss here crucial processes occurring at the plant cell wall during the contact and communication between microbe and plant. Finally, we argue that these local and dynamic changes need to be considered to fully understand plant-microbe interactions.
© The Author(s) 2022. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures
Similar articles
-
Between Host and Invaders: The Subcellular Cell Wall Dynamics at the Plant-Pathogen Interface.Annu Rev Plant Biol. 2025 May;76(1):255-284. doi: 10.1146/annurev-arplant-061824-115733. Annu Rev Plant Biol. 2025. PMID: 40393731 Review.
-
The Expression of Potato Expansin A3 (StEXPA3) and Extensin4 (StEXT4) Genes with Distribution of StEXPAs and HRGPs-Extensin Changes as an Effect of Cell Wall Rebuilding in Two Types of PVYNTN-Solanum tuberosum Interactions.Viruses. 2020 Jan 5;12(1):66. doi: 10.3390/v12010066. Viruses. 2020. PMID: 31948116 Free PMC article.
-
Post-Synthetic Defucosylation of AGP by Aspergillus nidulans α-1,2-Fucosidase Expressed in Arabidopsis Apoplast Induces Compensatory Upregulation of α-1,2-Fucosyltransferases.PLoS One. 2016 Jul 22;11(7):e0159757. doi: 10.1371/journal.pone.0159757. eCollection 2016. PLoS One. 2016. PMID: 27448235 Free PMC article.
-
Apoplast as the site of response to environmental signals.J Plant Res. 1998 Mar;111(1101):167-77. doi: 10.1007/BF02507163. J Plant Res. 1998. PMID: 11541948 Review.
-
Plant cell wall-mediated immunity: cell wall changes trigger disease resistance responses.Plant J. 2018 Feb;93(4):614-636. doi: 10.1111/tpj.13807. Epub 2018 Feb 2. Plant J. 2018. PMID: 29266460 Review.
Cited by
-
The root pathogen Aphanomyces euteiches secretes modular proteases in pea apoplast during host infection.Front Plant Sci. 2023 Mar 27;14:1140101. doi: 10.3389/fpls.2023.1140101. eCollection 2023. Front Plant Sci. 2023. PMID: 37051076 Free PMC article.
-
Roles of RNA silencing in viral and non-viral plant immunity and in the crosstalk between disease resistance systems.Nat Rev Mol Cell Biol. 2022 Oct;23(10):645-662. doi: 10.1038/s41580-022-00496-5. Epub 2022 Jun 16. Nat Rev Mol Cell Biol. 2022. PMID: 35710830 Review.
-
A legume cellulase required for rhizobial infection and colonization in root nodule symbiosis.Nat Commun. 2025 Jul 19;16(1):6663. doi: 10.1038/s41467-025-62083-4. Nat Commun. 2025. PMID: 40683892 Free PMC article.
-
Leaf microbiome dysbiosis triggered by T2SS-dependent enzyme secretion from opportunistic Xanthomonas pathogens.Nat Microbiol. 2024 Jan;9(1):136-149. doi: 10.1038/s41564-023-01555-z. Epub 2024 Jan 3. Nat Microbiol. 2024. PMID: 38172620 Free PMC article.
-
A peptide-receptor module links cell wall integrity sensing to pattern-triggered immunity.Nat Plants. 2024 Dec;10(12):2027-2037. doi: 10.1038/s41477-024-01840-7. Epub 2024 Oct 31. Nat Plants. 2024. PMID: 39482527
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
