

Fsp27 plays a crucial role in muscle performance
- PMID: 35157807
- PMCID: PMC8957325
- DOI: 10.1152/ajpendo.00255.2021
Fsp27 plays a crucial role in muscle performance
Abstract
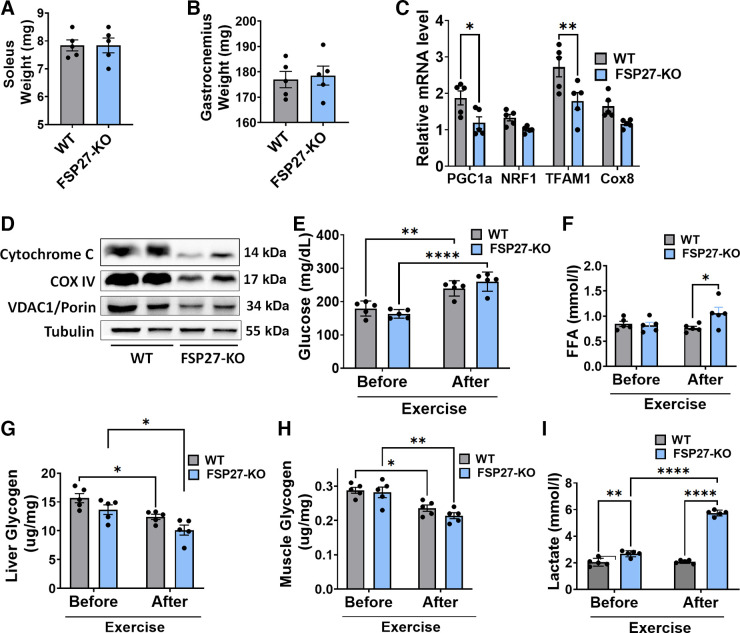
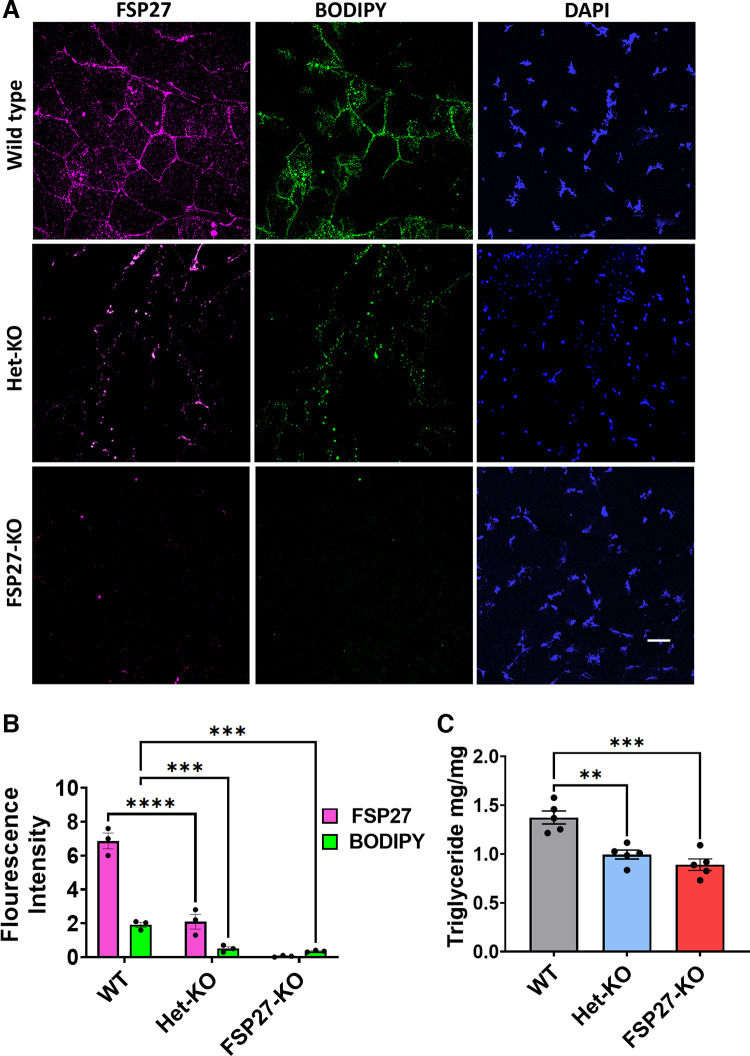
Fsp27 was previously identified as a lipid droplet-associated protein in adipocytes. Various studies have shown that it plays a role in the regulation of lipid homeostasis in adipose tissue and liver. However, its function in muscle, which also accumulate and metabolize fat, remains completely unknown. Our present study identifies a novel role of Fsp27 in muscle performance. Here, we demonstrate that Fsp27-/- and Fsp27+/- mice, both males and females, had severely impaired muscle endurance and exercise capacity compared with wild-type controls. Liver and muscle glycogen stores were similar among all groups fed or fasted, and before or after exercise. Reduced muscle performance in Fsp27-/- and Fsp27+/- mice was associated with severely decreased fat content in the muscle. Furthermore, results in heterozygous Fsp27+/- mice indicate that Fsp27 haploinsufficiency undermines muscle performance in both males and females. In summary, our physiological findings reveal that Fsp27 plays a critical role in muscular fat storage, muscle endurance, and muscle strength.NEW & NOTEWORTHY This is the first study identifying Fsp27 as a novel protein associated with muscle metabolism. The Fsp27-knockout model shows that Fsp27 plays a role in muscular-fat storage, muscle endurance, and muscle strength, which ultimately impacts limb movement. In addition, our study suggests a potential metabolic paradox in which FSP27-knockout mice presumed to be metabolically healthy based on glucose utilization and oxidative metabolism are unhealthy in terms of exercise capacity and muscular performance.
Keywords: Cidea; Cidec; diabetes; fat metabolism; lipid droplets; lipids; obesity.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures







References
-
- Grahn THM, Kaur R, Yin J, Schweiger M, Sharma VM, Lee MJ, Ido Y, Smas CM, Zechner R, Lass A, Puri V. Fat-specific protein 27 (FSP27) interacts with adipose triglyceride lipase (ATGL) to regulate lipolysis and insulin sensitivity in human adipocytes. J Biol Chem 289: 12029–12039, 2014. doi: 10.1074/jbc.M113.539890. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases