

Induction of Survival of Motor Neuron (SMN) Protein Deficiency in Spinal Astrocytes by Small Interfering RNA as an In Vitro Model of Spinal Muscular Atrophy
- PMID: 35159369
- PMCID: PMC8834238
- DOI: 10.3390/cells11030558
Induction of Survival of Motor Neuron (SMN) Protein Deficiency in Spinal Astrocytes by Small Interfering RNA as an In Vitro Model of Spinal Muscular Atrophy
Abstract
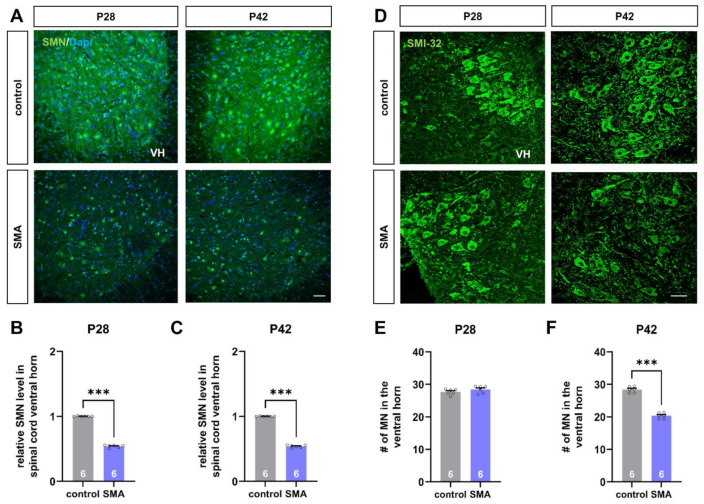
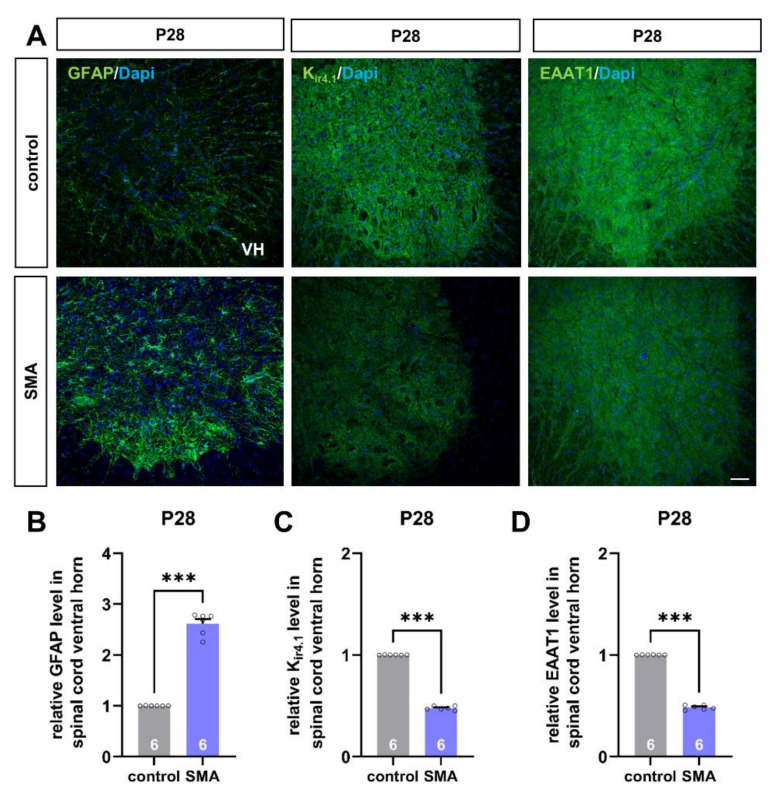
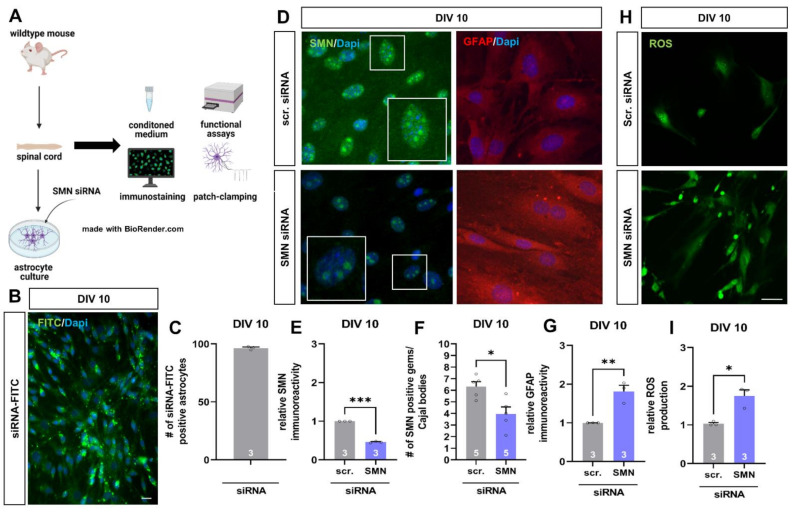
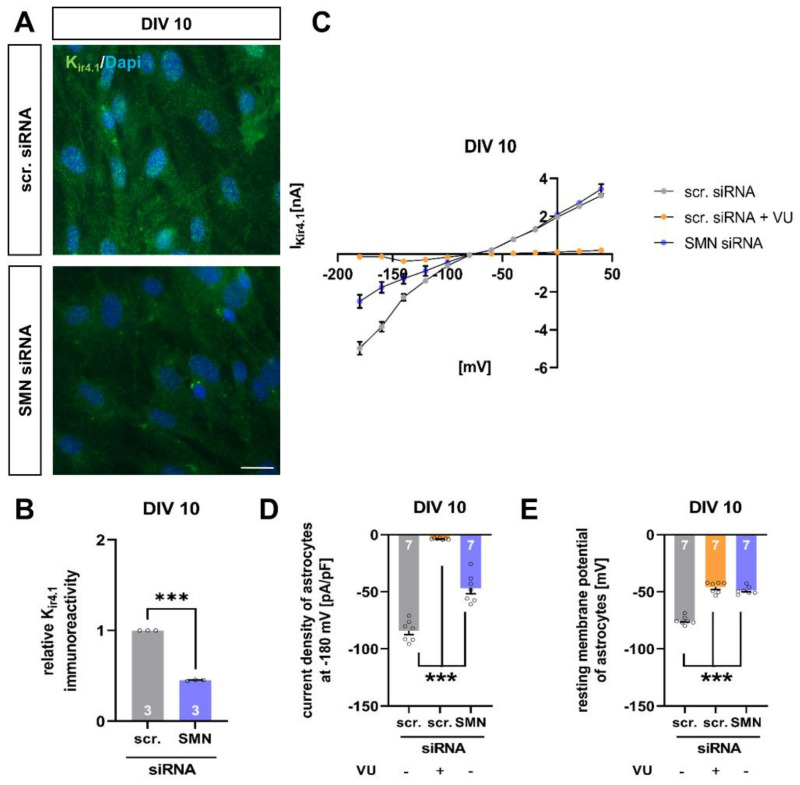
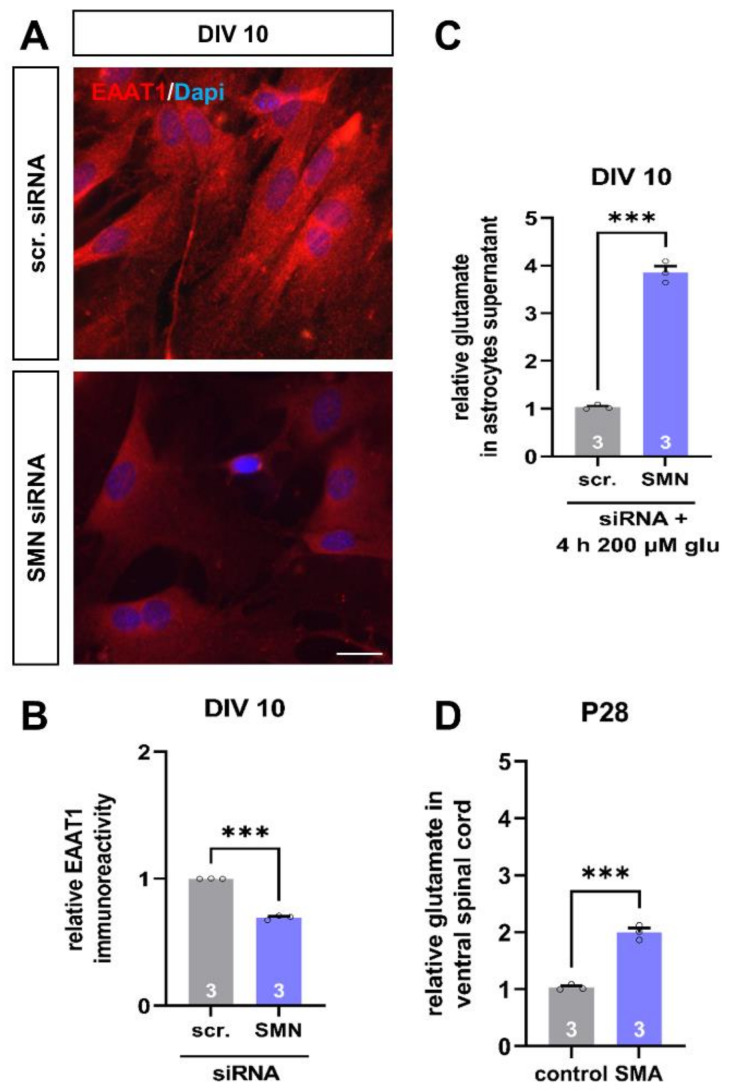
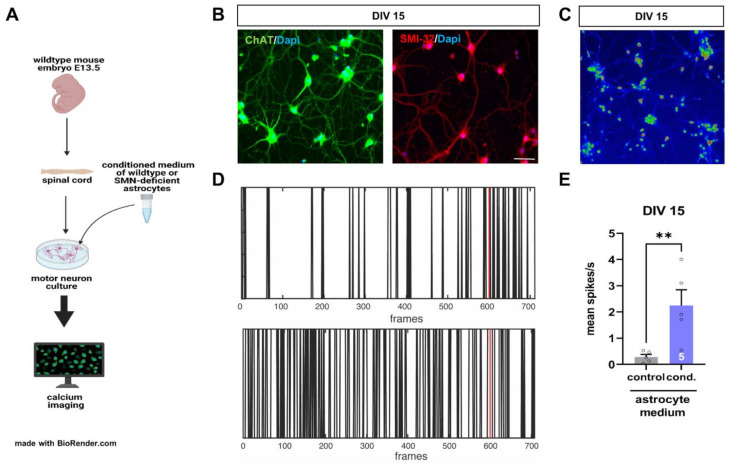
Spinal muscular atrophy (SMA) is a motor neuron disorder leading to progressive loss of ventral horn neurons resulting in muscle wasting. Here we investigate the contribution of spinal astrocytes to the pathogenesis of late-onset SMA forms using a mouse model. Furthermore, we generated SMA-like astrocytes using survival of motor neuron (SMN) siRNA transfection techniques. In the SMA mouse model, the activation of spinal astrocytes and the reduction of the inward rectifier potassium channel Kir4.1 and excitatory amino acid transporter 1 (EAAT1) were observed at postnatal day (P) 28, preceding the loss of spinal motor neurons appearing earliest at P42. Using SMA-like astrocytes, we could mimic the modulation of spinal astrocytes of the mouse model in a dish and perform electrophysiological assessments and functional assays. In SMA-like astrocytes, glutamate uptake was diminished due to a reduction in EAAT1. Furthermore, patch-clamp measurements revealed reduced potassium uptake into astrocytes with membrane depolarization. Additionally, exposure of healthy spinal motor neurons to a conditioned medium of SMA-like astrocytes resulted in increased firing frequency. These data demonstrate spinal astrocytes' crucial role in the late-onset SMA forms' pathogenesis.
Keywords: SMA; SMN; astrocytes; glia–neuron interaction; neuromuscular disorders; siRNA; spinal cord; spinal muscular atrophy; survival of motor neuron.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Spinal astrocyte dysfunction drives motor neuron loss in late-onset spinal muscular atrophy.Acta Neuropathol. 2023 May;145(5):611-635. doi: 10.1007/s00401-023-02554-4. Epub 2023 Mar 17. Acta Neuropathol. 2023. PMID: 36930296 Free PMC article.
-
Diminished motor neuron activity driven by abnormal astrocytic EAAT1 glutamate transporter activity in spinal muscular atrophy is not fully restored after lentiviral SMN delivery.Glia. 2023 May;71(5):1311-1332. doi: 10.1002/glia.24340. Epub 2023 Jan 18. Glia. 2023. PMID: 36655314
-
Spinal motor neuron loss occurs through a p53-and-p21-independent mechanism in the Smn2B/- mouse model of spinal muscular atrophy.Exp Neurol. 2021 Mar;337:113587. doi: 10.1016/j.expneurol.2020.113587. Epub 2020 Dec 28. Exp Neurol. 2021. PMID: 33382987 Free PMC article.
-
Spinal muscular atrophy: a motor neuron disorder or a multi-organ disease?J Anat. 2014 Jan;224(1):15-28. doi: 10.1111/joa.12083. Epub 2013 Jul 22. J Anat. 2014. PMID: 23876144 Free PMC article. Review.
-
Glial cells involvement in spinal muscular atrophy: Could SMA be a neuroinflammatory disease?Neurobiol Dis. 2020 Jul;140:104870. doi: 10.1016/j.nbd.2020.104870. Epub 2020 Apr 12. Neurobiol Dis. 2020. PMID: 32294521 Review.
Cited by
-
Spinal astrocyte dysfunction drives motor neuron loss in late-onset spinal muscular atrophy.Acta Neuropathol. 2023 May;145(5):611-635. doi: 10.1007/s00401-023-02554-4. Epub 2023 Mar 17. Acta Neuropathol. 2023. PMID: 36930296 Free PMC article.
-
Glial Cells in Spinal Muscular Atrophy: Speculations on Non-Cell-Autonomous Mechanisms and Therapeutic Implications.Neurol Int. 2025 Mar 13;17(3):41. doi: 10.3390/neurolint17030041. Neurol Int. 2025. PMID: 40137462 Free PMC article. Review.
-
Dysregulated balance of D- and L-amino acids modulating glutamatergic neurotransmission in severe spinal muscular atrophy.bioRxiv [Preprint]. 2024 Oct 22:2024.10.22.619645. doi: 10.1101/2024.10.22.619645. bioRxiv. 2024. Update in: Neurobiol Dis. 2025 Apr;207:106849. doi: 10.1016/j.nbd.2025.106849. PMID: 39484528 Free PMC article. Updated. Preprint.
-
Dysregulated balance of D- and L-amino acids modulating glutamatergic neurotransmission in severe spinal muscular atrophy.Neurobiol Dis. 2025 Apr;207:106849. doi: 10.1016/j.nbd.2025.106849. Epub 2025 Feb 24. Neurobiol Dis. 2025. PMID: 40010612 Free PMC article.
-
Neuronal Pentraxin 2 as a Potential Biomarker for Nusinersen Therapy Response in Adults with Spinal Muscular Atrophy: A Pilot Study.Biomedicines. 2025 Jul 25;13(8):1821. doi: 10.3390/biomedicines13081821. Biomedicines. 2025. PMID: 40868076 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
