

The CAR-mRNA Interaction Surface Is a Zipper Extension of the Ribosome A Site
- PMID: 35163343
- PMCID: PMC8835751
- DOI: 10.3390/ijms23031417
The CAR-mRNA Interaction Surface Is a Zipper Extension of the Ribosome A Site
Abstract
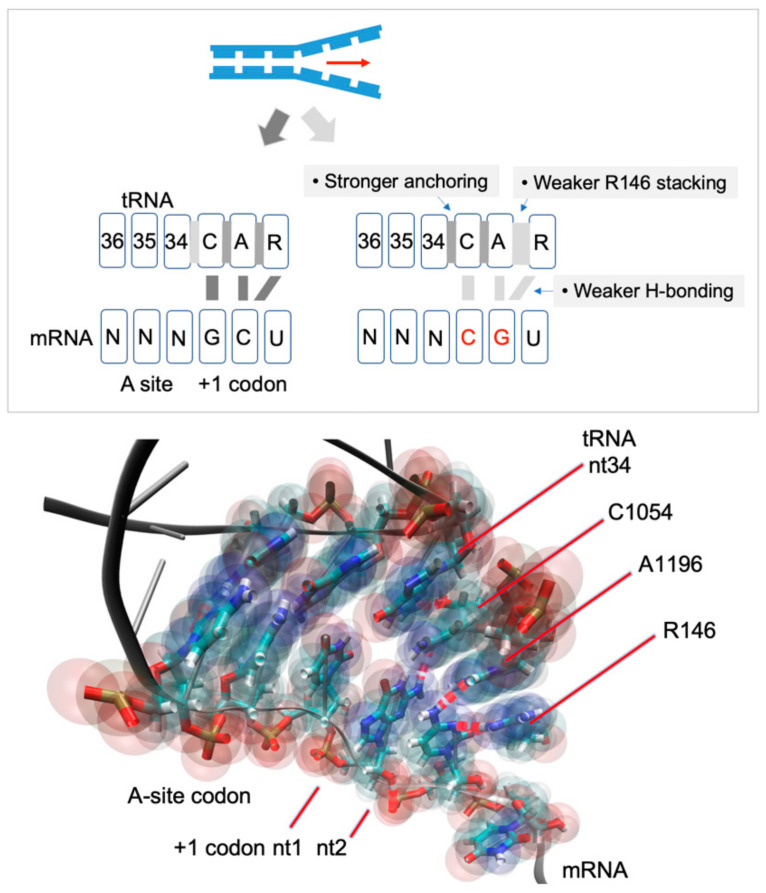
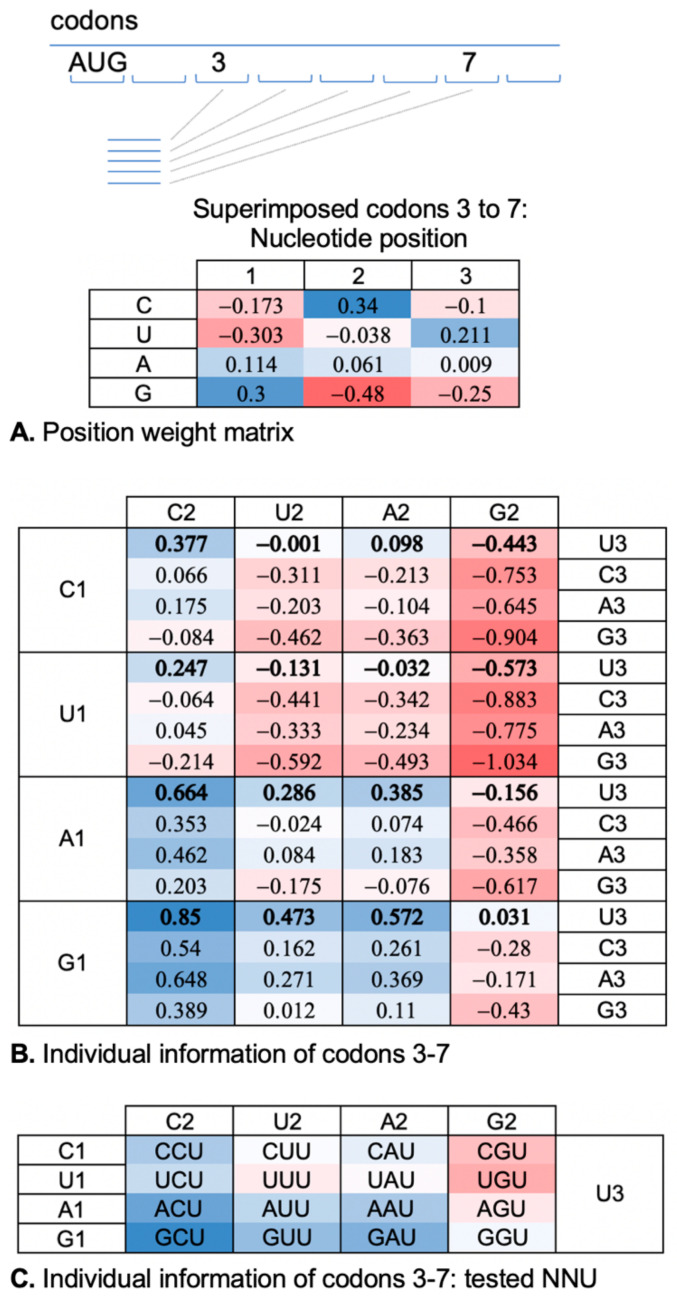
The ribosome CAR interaction surface behaves as an extension of the decoding center A site and has H-bond interactions with the +1 codon, which is next in line to enter the A site. Through molecular dynamic simulations, we investigated the codon sequence specificity of this CAR-mRNA interaction and discovered a strong preference for GCN codons, suggesting that there may be a sequence-dependent layer of translational regulation dependent on the CAR interaction surface. Dissection of the CAR-mRNA interaction through nucleotide substitution experiments showed that the first nucleotide of the +1 codon dominates over the second nucleotide position, consistent with an energetically favorable zipper-like activity that emanates from the A site through the CAR-mRNA interface. Moreover, the CAR/+1 codon interaction is affected by the identity of nucleotide 3 of +1 GCN codons, which influences the stacking of G and C. Clustering analysis suggests that the A-site decoding center adopts different neighborhood substates that depend on the identity of the +1 codon.
Keywords: A-site decoding center; CAR interaction surface; molecular dynamics; ribosome substates; ribosome translocation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
