

Kidney Injury Causes Accumulation of Renal Sodium That Modulates Renal Lymphatic Dynamics
- PMID: 35163352
- PMCID: PMC8836121
- DOI: 10.3390/ijms23031428
Kidney Injury Causes Accumulation of Renal Sodium That Modulates Renal Lymphatic Dynamics
Abstract
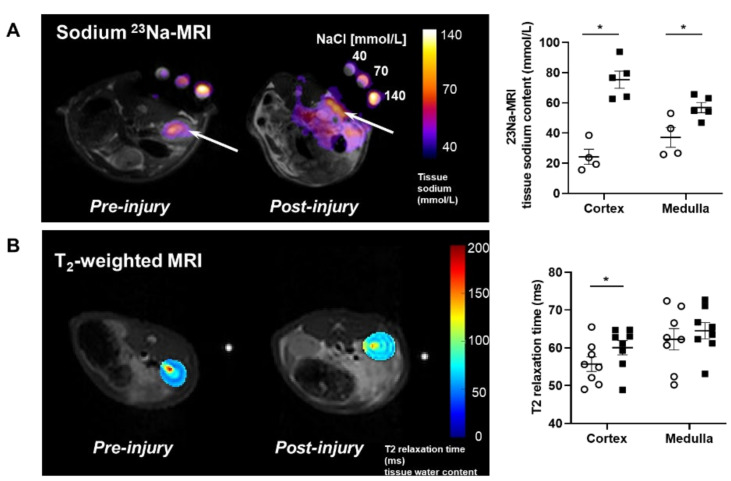
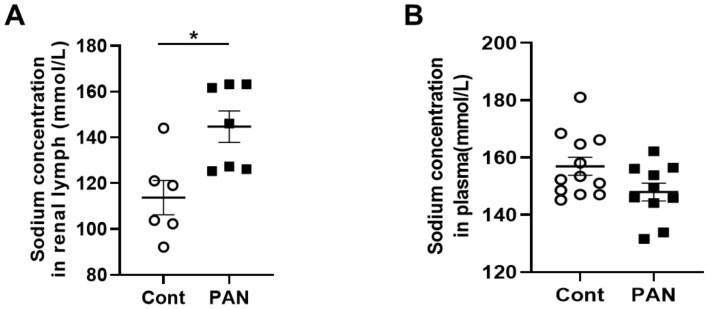
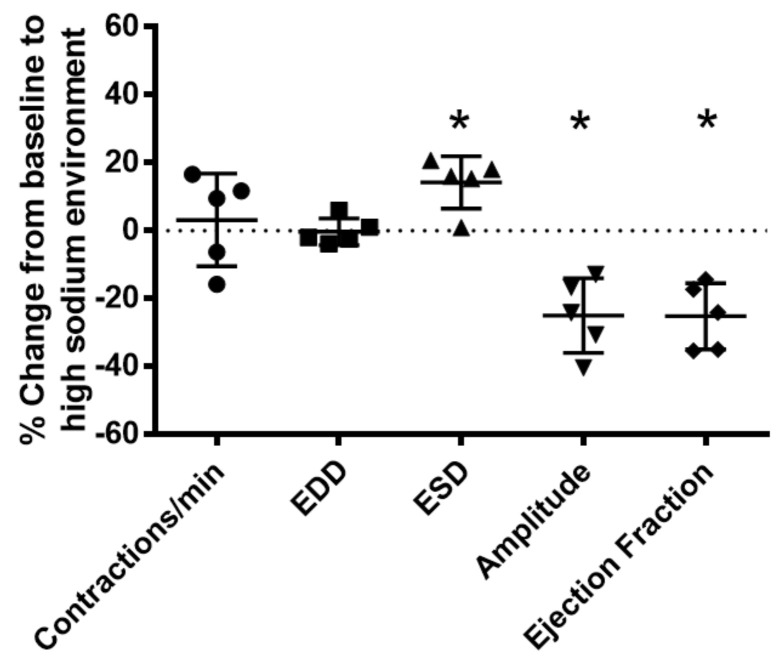
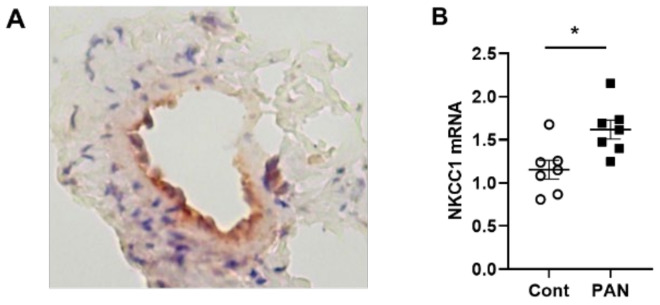
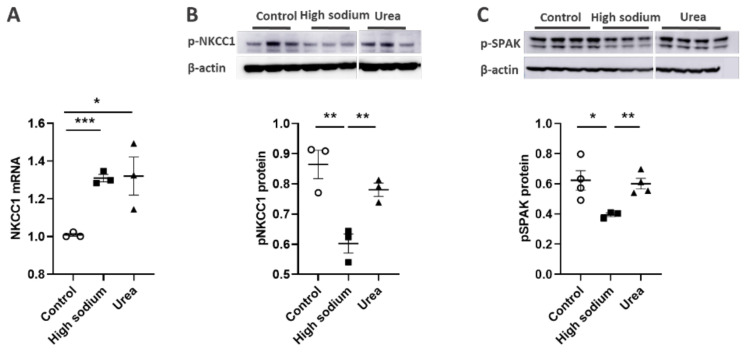
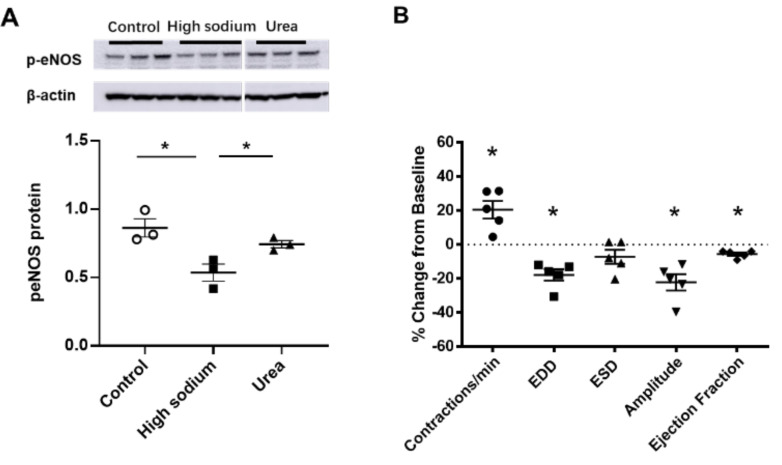
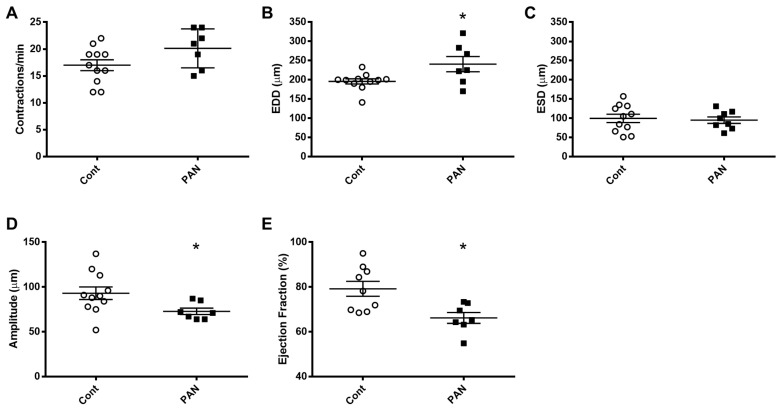
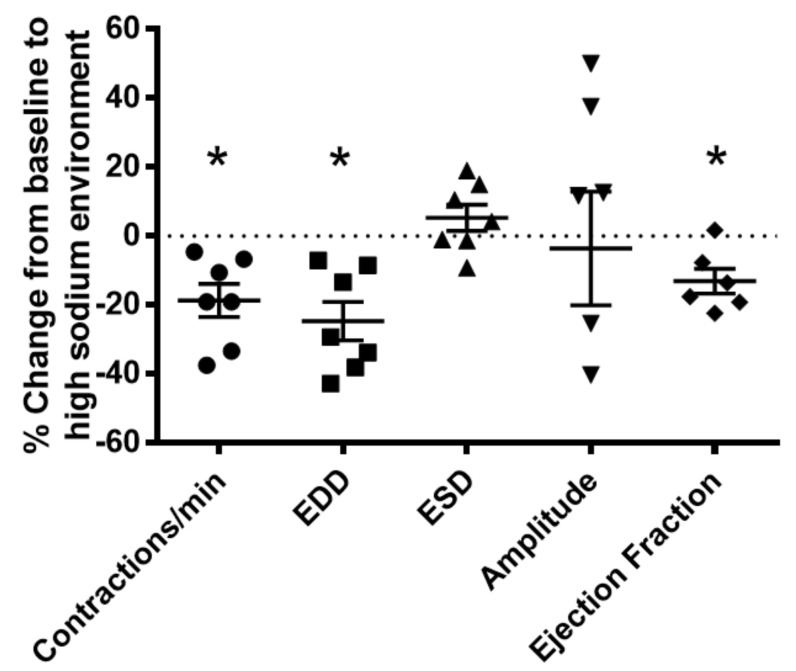
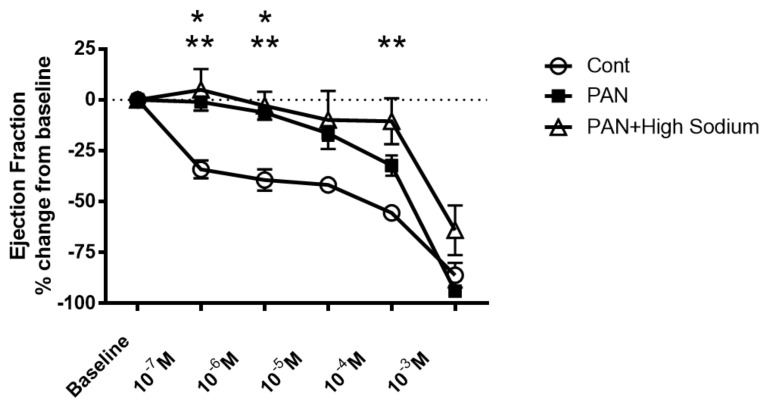
Lymphatic vessels are highly responsive to changes in the interstitial environment. Previously, we showed renal lymphatics express the Na-K-2Cl cotransporter. Since interstitial sodium retention is a hallmark of proteinuric injury, we examined whether renal sodium affects NKCC1 expression and the dynamic pumping function of renal lymphatic vessels. Puromycin aminonucleoside (PAN)-injected rats served as a model of proteinuric kidney injury. Sodium 23Na/1H-MRI was used to measure renal sodium and water content in live animals. Renal lymph, which reflects the interstitial composition, was collected, and the sodium analyzed. The contractile dynamics of isolated renal lymphatic vessels were studied in a perfusion chamber. Cultured lymphatic endothelial cells (LECs) were used to assess direct sodium effects on NKCC1. MRI showed elevation in renal sodium and water in PAN. In addition, renal lymph contained higher sodium, although the plasma sodium showed no difference between PAN and controls. High sodium decreased contractility of renal collecting lymphatic vessels. In LECs, high sodium reduced phosphorylated NKCC1 and SPAK, an upstream activating kinase of NKCC1, and eNOS, a downstream effector of lymphatic contractility. The NKCC1 inhibitor furosemide showed a weaker effect on ejection fraction in isolated renal lymphatics of PAN vs controls. High sodium within the renal interstitium following proteinuric injury is associated with impaired renal lymphatic pumping that may, in part, involve the SPAK-NKCC1-eNOS pathway, which may contribute to sodium retention and reduce lymphatic responsiveness to furosemide. We propose that this lymphatic vessel dysfunction is a novel mechanism of impaired interstitial clearance and edema in proteinuric kidney disease.
Keywords: NKCC1 transporter; kidney; lymphatics; sodium.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
