

Role of ammonia for brain abnormal protein glycosylation during the development of hepatitis B virus-related liver diseases
- PMID: 35164881
- PMCID: PMC8842931
- DOI: 10.1186/s13578-022-00751-4
Role of ammonia for brain abnormal protein glycosylation during the development of hepatitis B virus-related liver diseases
Abstract
Background: Ammonia is the most typical neurotoxin in hepatic encephalopathy (HE), but the underlying pathophysiology between ammonia and aberrant glycosylation in HE remains unknown.
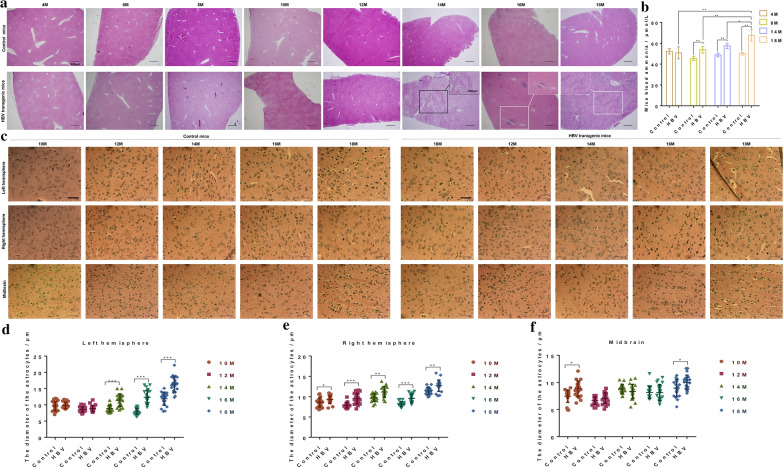
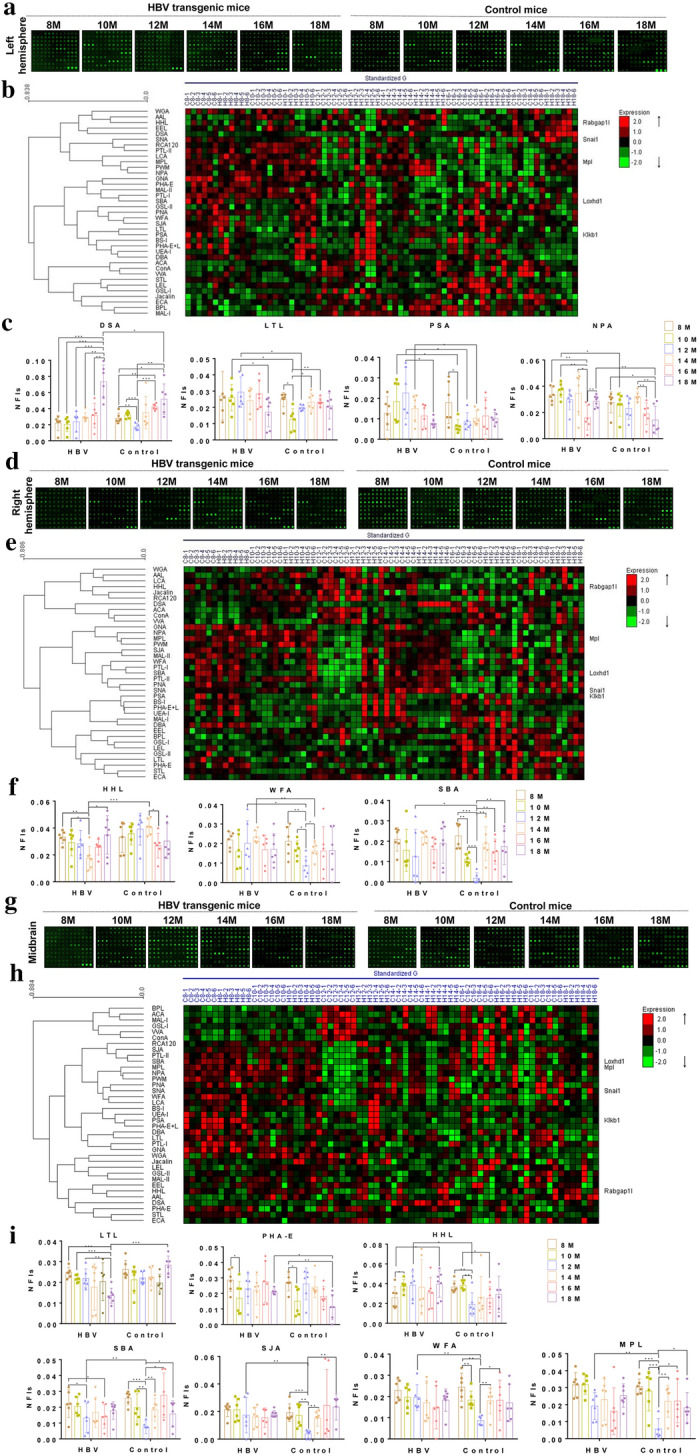
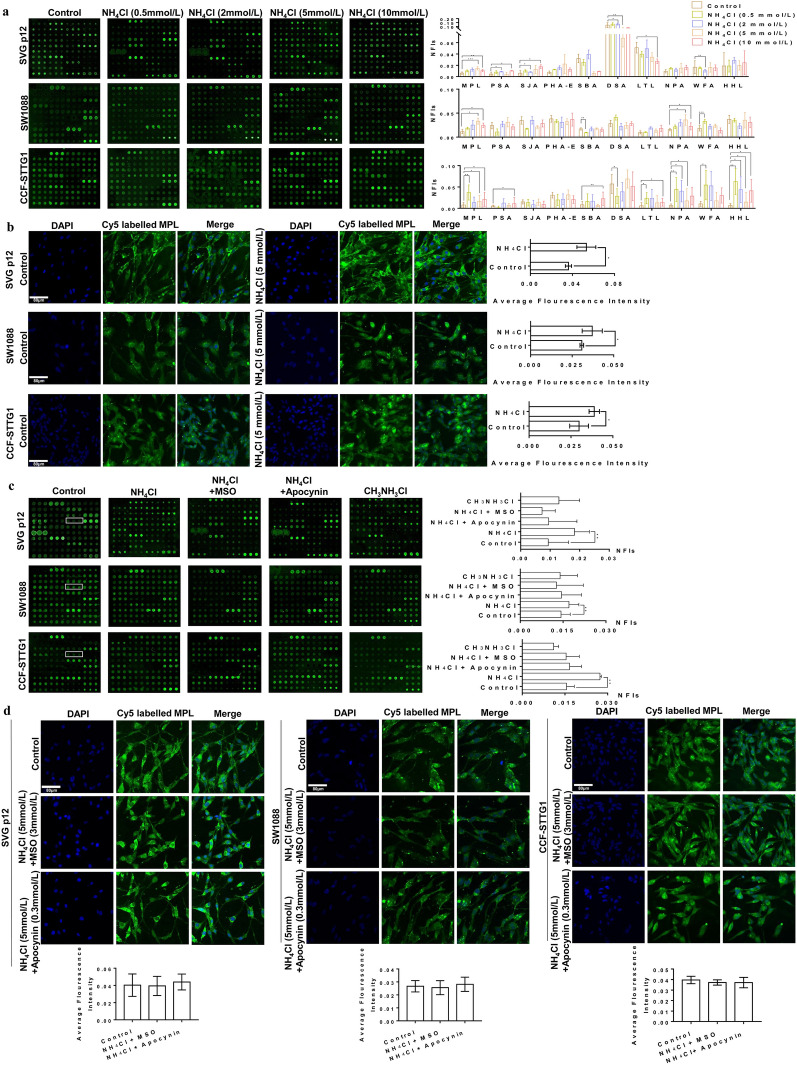
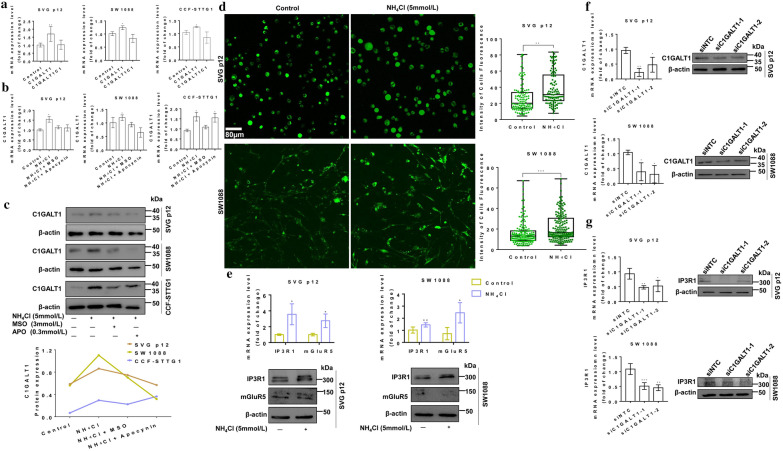
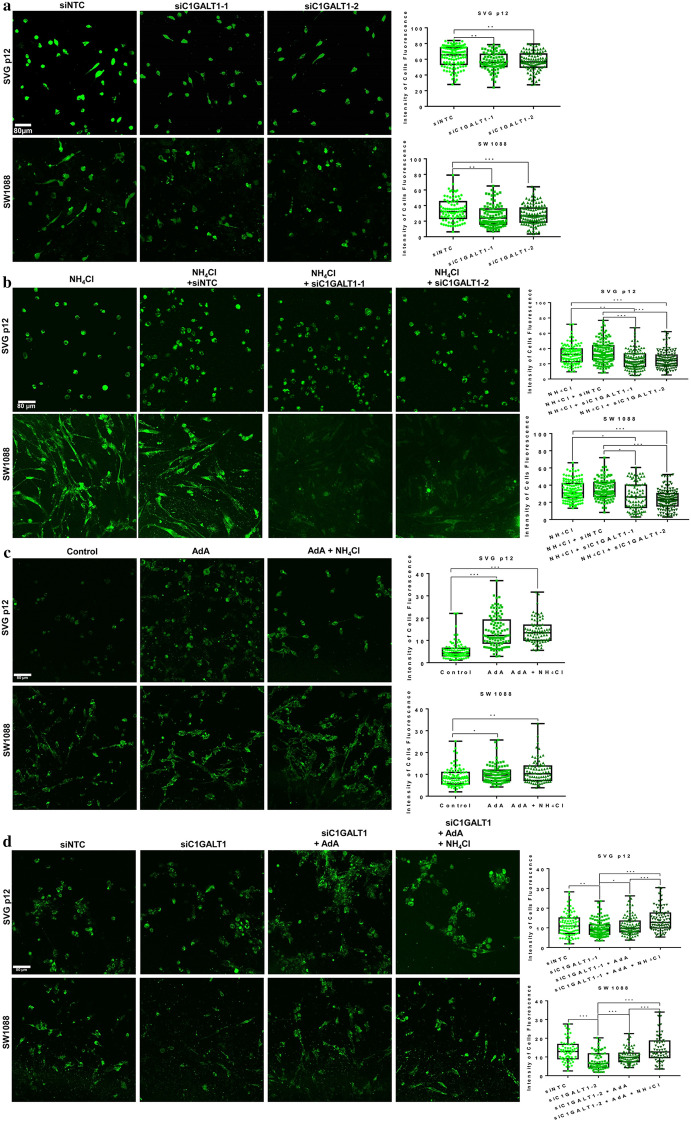
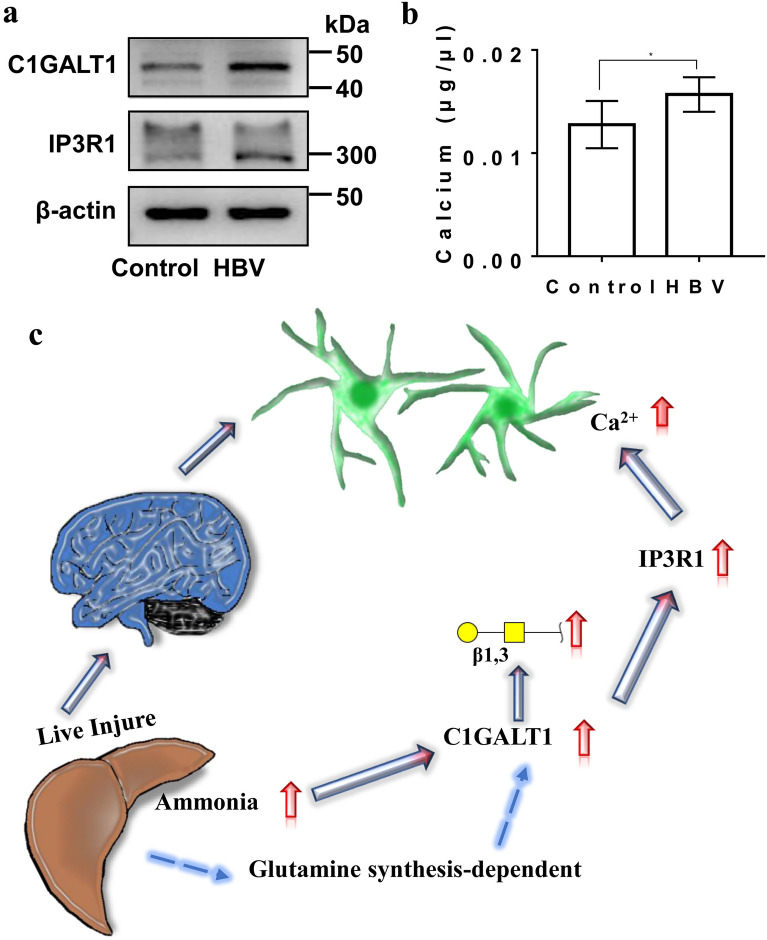
Results: Here, we used HBV transgenic mice and astrocytes to present a systems-based study of glycosylation changes and corresponding enzymes associated with the key factors of ammonia in HE. We surveyed protein glycosylation changes associated with the brain of HBV transgenic mice by lectin microarrays. Upregulation of Galβ1-3GalNAc mediated by core 1 β1,3-galactosyltransferase (C1GALT1) was identified as a result of ammonia stimulation. Using in vitro assays, we validated that upregulation of C1GALT1 is a driver of deregulates calcium (Ca2+) homeostasis by overexpression of inositol 1,4,5-trisphosphate receptor type 1 (IP3R1) in astrocytes.
Conclusions: We demonstrated that silencing C1GALT1 could depress the IP3R1 expression, an effective strategy to inhibit the ammonia-induced upregulation of Ca2+ activity, thereby C1GALT1 and IP3R1 may serve as therapeutic targets in hyperammonemia of HE.
Keywords: C1GALT1; Ca2+ homeostasis; Hepatic encephalopathy; Hyperammonemia; IP3R1.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
C1GALT1 overexpression promotes the invasive behavior of colon cancer cells through modifying O-glycosylation of FGFR2.Oncotarget. 2014 Apr 30;5(8):2096-106. doi: 10.18632/oncotarget.1815. Oncotarget. 2014. PMID: 24758762 Free PMC article.
-
C1GALT1 in health and disease.Oncol Lett. 2021 Aug;22(2):589. doi: 10.3892/ol.2021.12850. Epub 2021 Jun 6. Oncol Lett. 2021. PMID: 34149900 Free PMC article. Review.
-
Hepatic encephalopathy is linked to alterations of autophagic flux in astrocytes.EBioMedicine. 2019 Oct;48:539-553. doi: 10.1016/j.ebiom.2019.09.058. Epub 2019 Oct 21. EBioMedicine. 2019. PMID: 31648987 Free PMC article.
-
O-GlcNAcylation-dependent upregulation of HO1 triggers ammonia-induced oxidative stress and senescence in hepatic encephalopathy.J Hepatol. 2019 Nov;71(5):930-941. doi: 10.1016/j.jhep.2019.06.020. Epub 2019 Jul 4. J Hepatol. 2019. PMID: 31279900
-
Update on the role of C1GALT1 in cancer.Oncol Lett. 2022 Mar;23(3):97. doi: 10.3892/ol.2022.13217. Epub 2022 Jan 27. Oncol Lett. 2022. PMID: 35154428 Free PMC article. Review.
Cited by
-
Cellular Pathogenesis of Hepatic Encephalopathy: An Update.Biomolecules. 2023 Feb 19;13(2):396. doi: 10.3390/biom13020396. Biomolecules. 2023. PMID: 36830765 Free PMC article. Review.
-
Regulation of adult neurogenesis: the crucial role of astrocytic mitochondria.Front Mol Neurosci. 2024 Nov 22;17:1516119. doi: 10.3389/fnmol.2024.1516119. eCollection 2024. Front Mol Neurosci. 2024. PMID: 39649104 Free PMC article. Review.
-
Antidiabetic activity of Tartary buckwheat protein-derived peptide AFYRW and its effects on protein glycosylation of pancreas in mice.Amino Acids. 2023 Aug;55(8):1063-1071. doi: 10.1007/s00726-023-03294-1. Epub 2023 Jun 21. Amino Acids. 2023. PMID: 37341830
-
Inhibition of Astrocytic Carbohydrate Sulfotransferase 15 Promotes Nerve Repair After Spinal Cord Injury via Mitigation of CSPG Mediated Axonal Inhibition.Cell Mol Neurobiol. 2023 Aug;43(6):2925-2937. doi: 10.1007/s10571-023-01333-9. Epub 2023 Mar 13. Cell Mol Neurobiol. 2023. PMID: 36913114 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
