

Widespread occurrence of covalent lysine-cysteine redox switches in proteins
- PMID: 35165445
- PMCID: PMC8964421
- DOI: 10.1038/s41589-021-00966-5
Widespread occurrence of covalent lysine-cysteine redox switches in proteins
Abstract
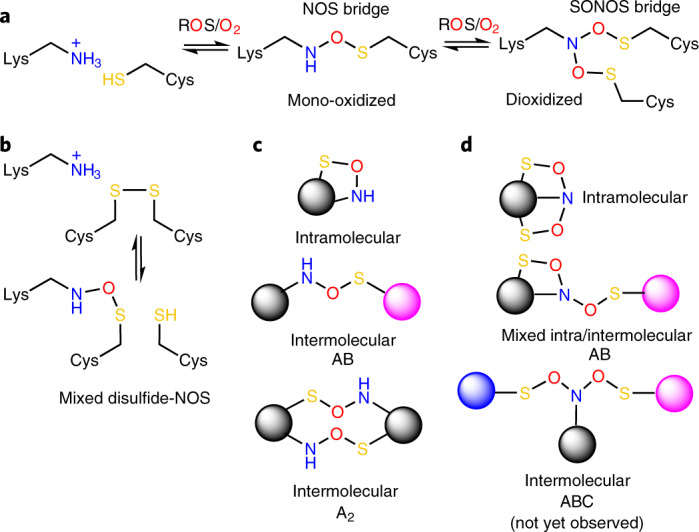
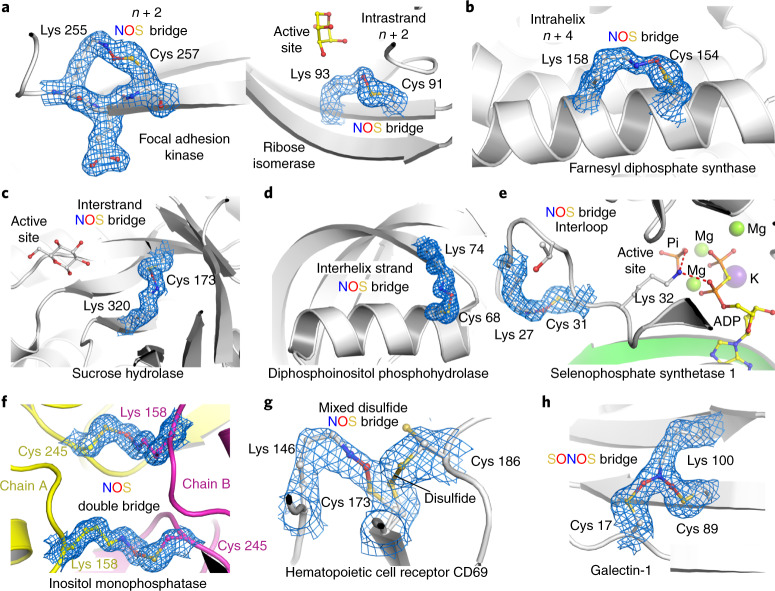
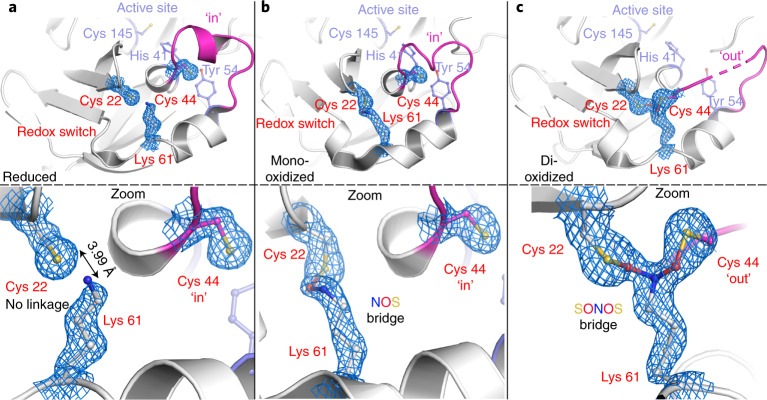
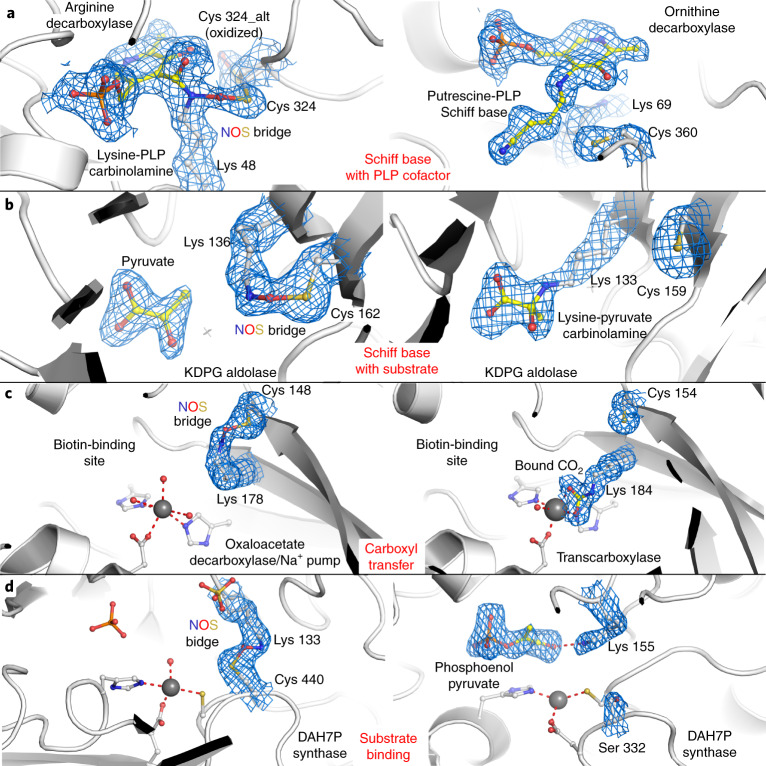
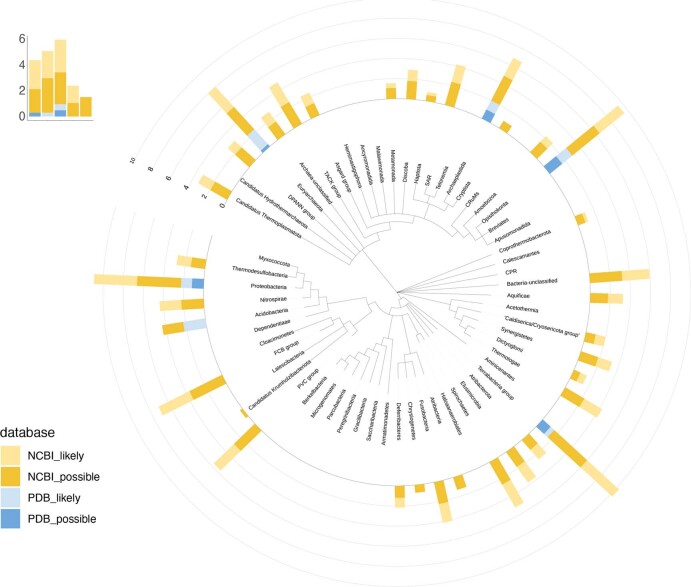
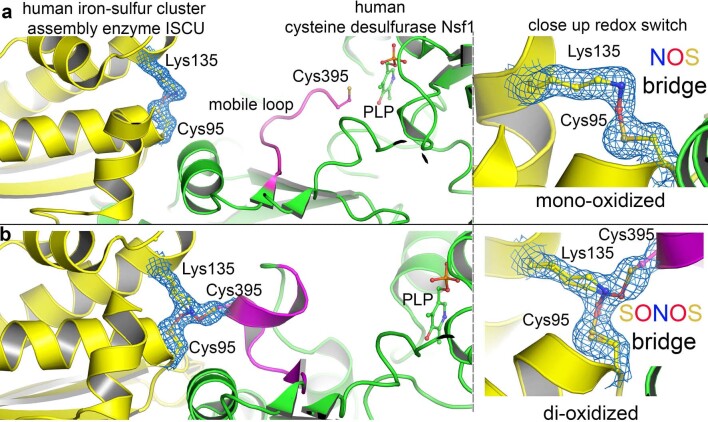
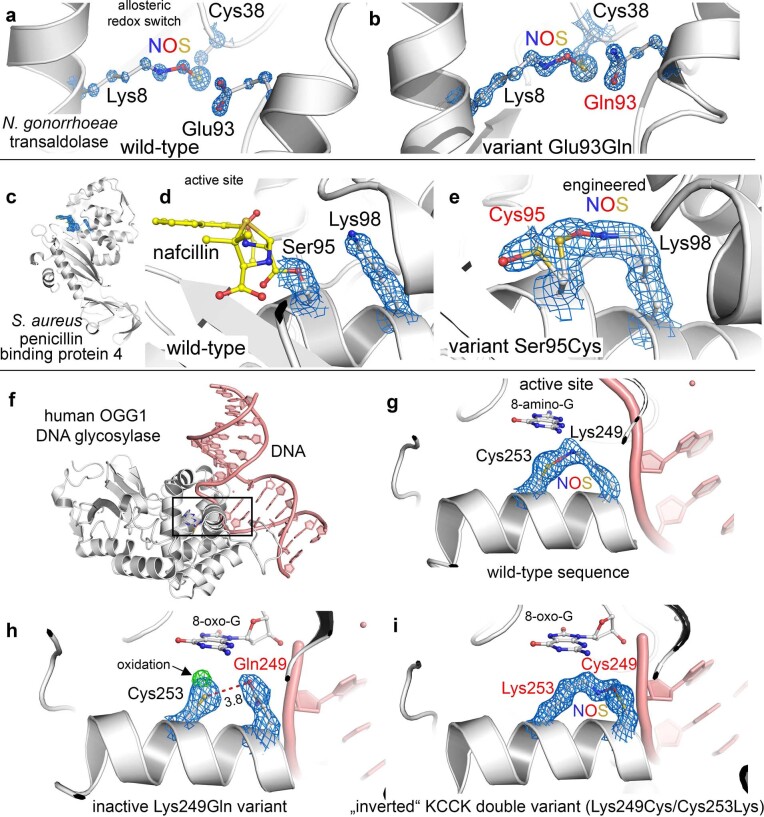
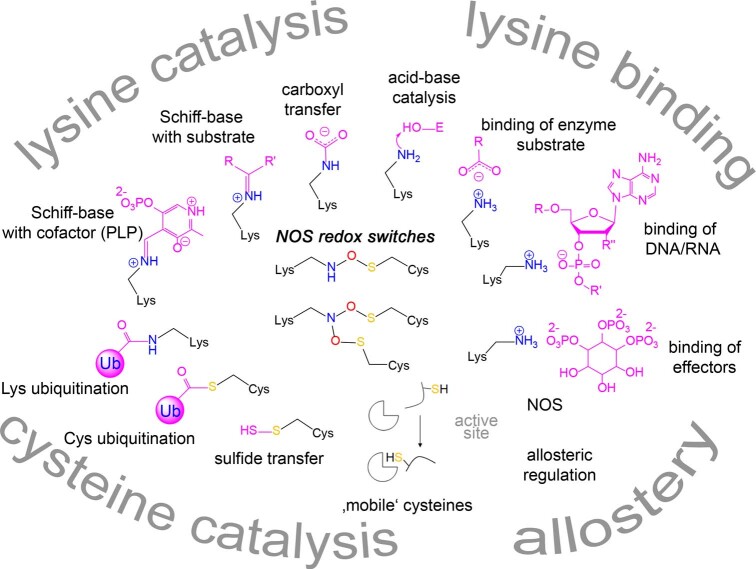
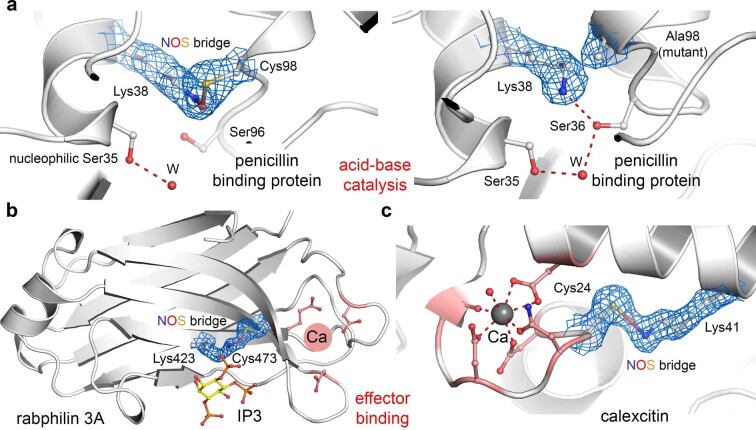
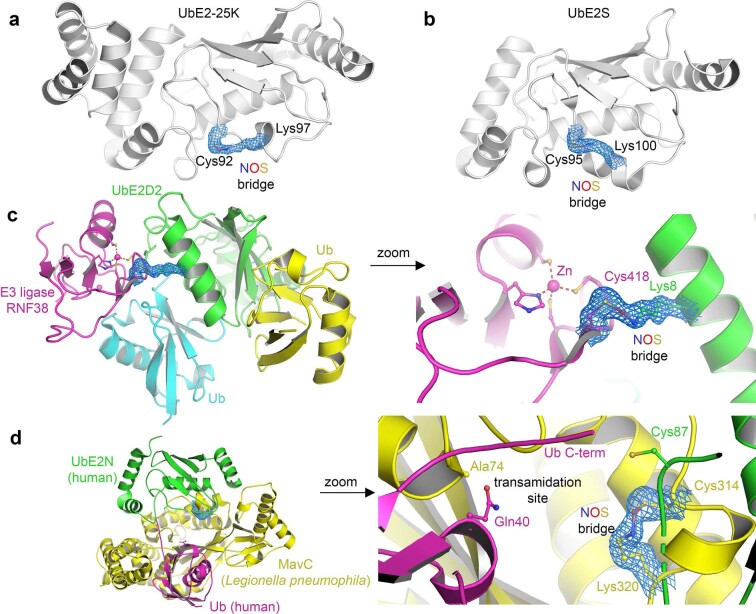
We recently reported the discovery of a lysine-cysteine redox switch in proteins with a covalent nitrogen-oxygen-sulfur (NOS) bridge. Here, a systematic survey of the whole protein structure database discloses that NOS bridges are ubiquitous redox switches in proteins of all domains of life and are found in diverse structural motifs and chemical variants. In several instances, lysines are observed in simultaneous linkage with two cysteines, forming a sulfur-oxygen-nitrogen-oxygen-sulfur (SONOS) bridge with a trivalent nitrogen, which constitutes an unusual native branching cross-link. In many proteins, the NOS switch contains a functionally essential lysine with direct roles in enzyme catalysis or binding of substrates, DNA or effectors, linking lysine chemistry and redox biology as a regulatory principle. NOS/SONOS switches are frequently found in proteins from human and plant pathogens, including severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), and also in many human proteins with established roles in gene expression, redox signaling and homeostasis in physiological and pathophysiological conditions.
© 2022. The Author(s).
Conflict of interest statement
F.R.v.P., M.W., L.-M.F., J.U., R.A.M. and K.T. have filed a European patent application (application number EP21164101.4) for regulating protein activities by targeting the NOS or SONOS redox switches. The remaining authors declare no competing interests.
Figures
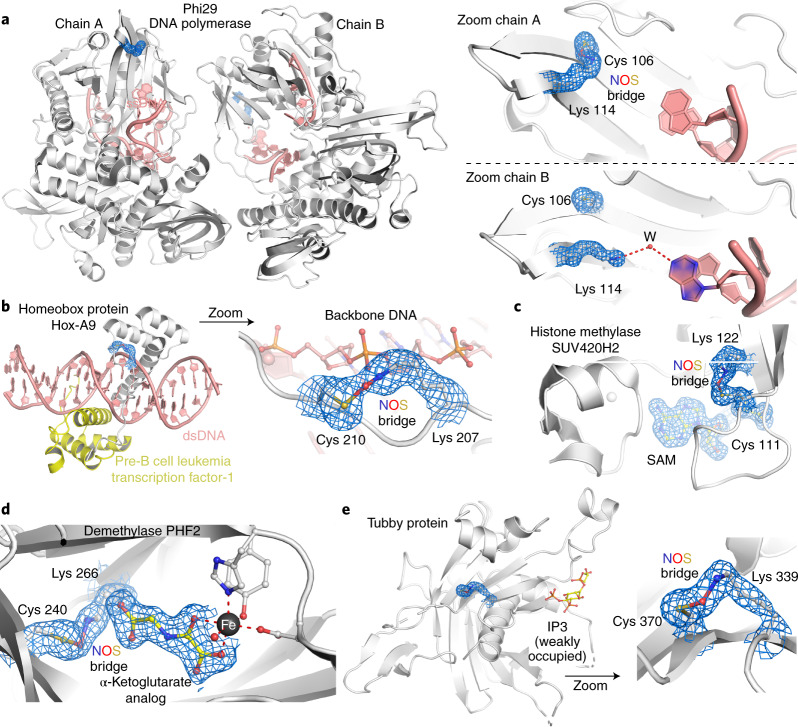
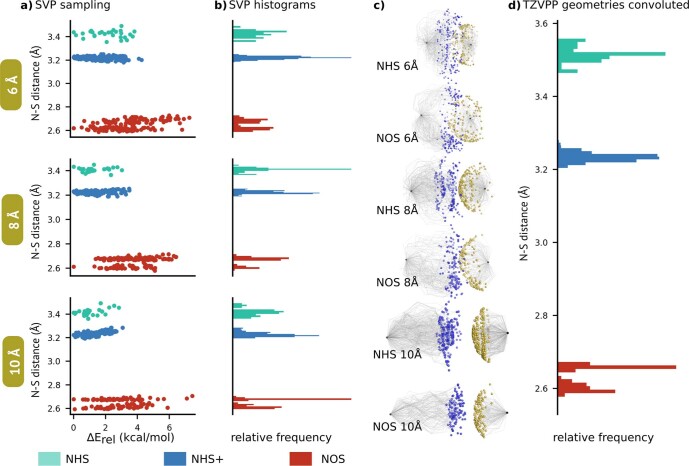
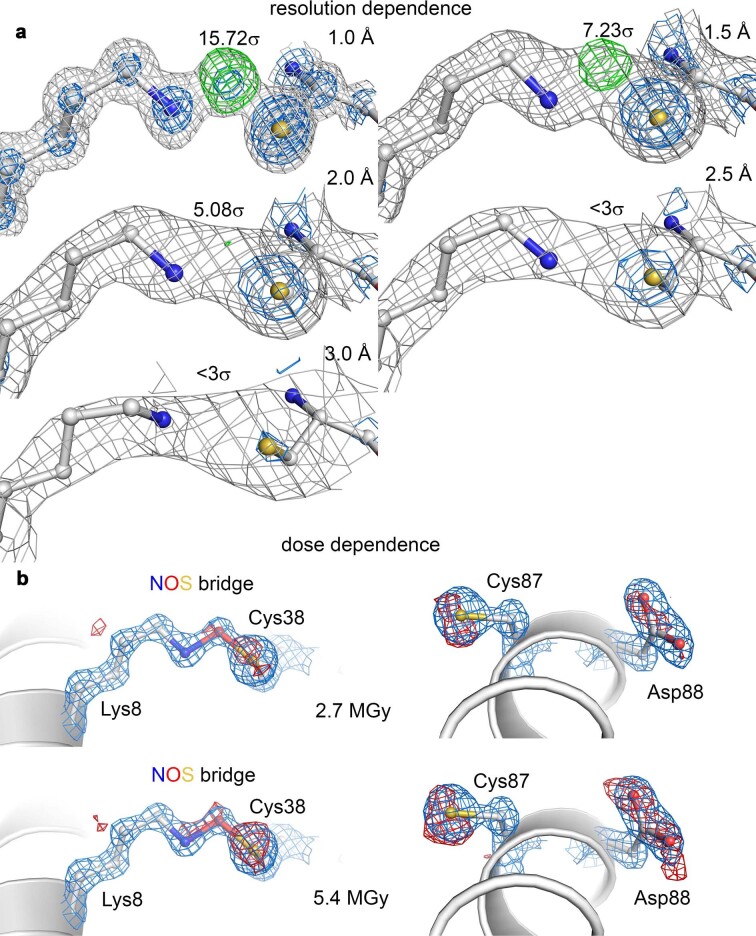
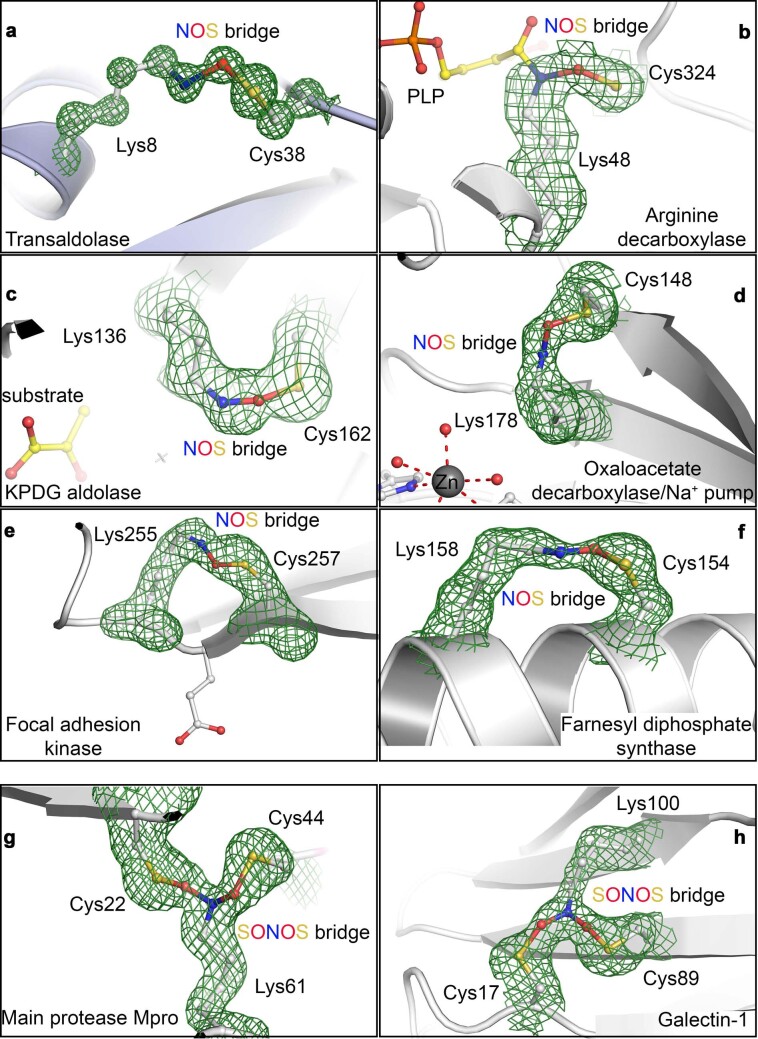
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
