

Flavonoids improve the stability and function of P23H rhodopsin slowing down the progression of retinitis pigmentosa in mice
- PMID: 35165923
- PMCID: PMC9615108
- DOI: 10.1002/jnr.25021
Flavonoids improve the stability and function of P23H rhodopsin slowing down the progression of retinitis pigmentosa in mice
Abstract
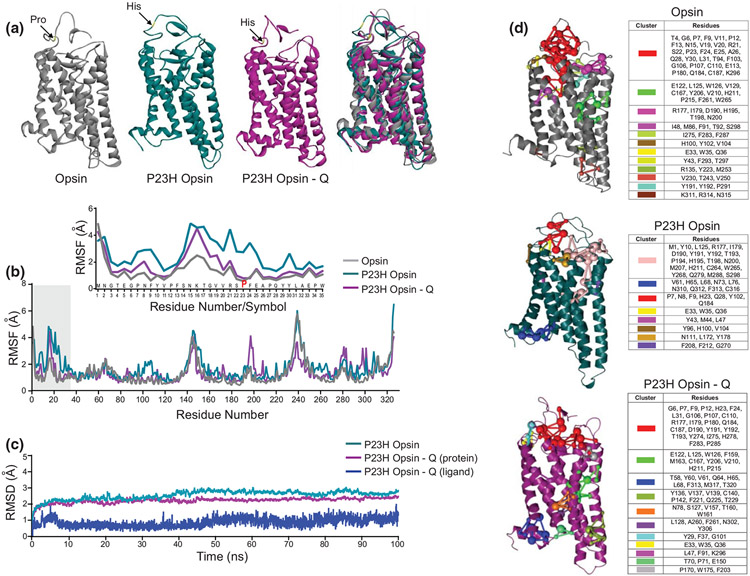
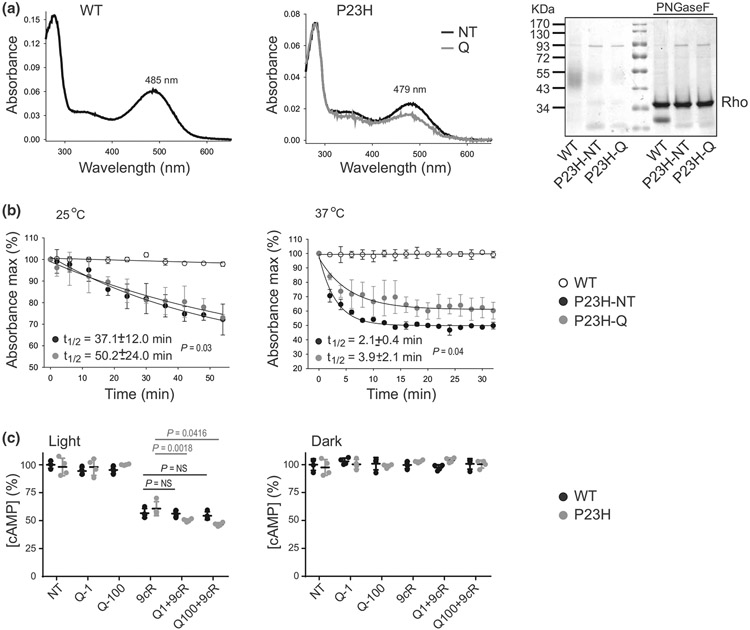
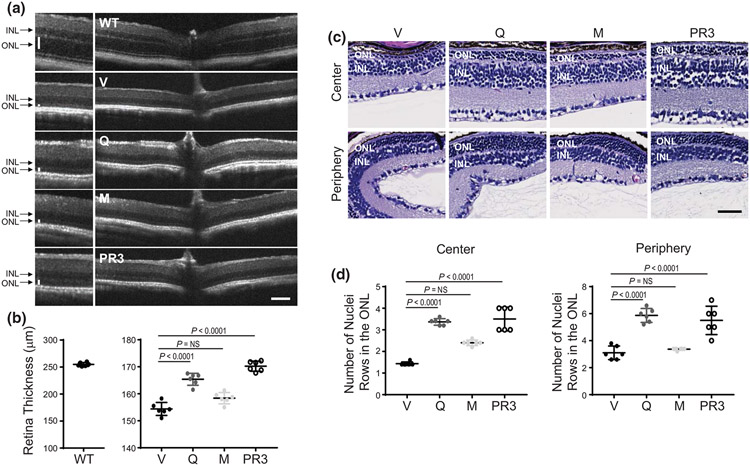
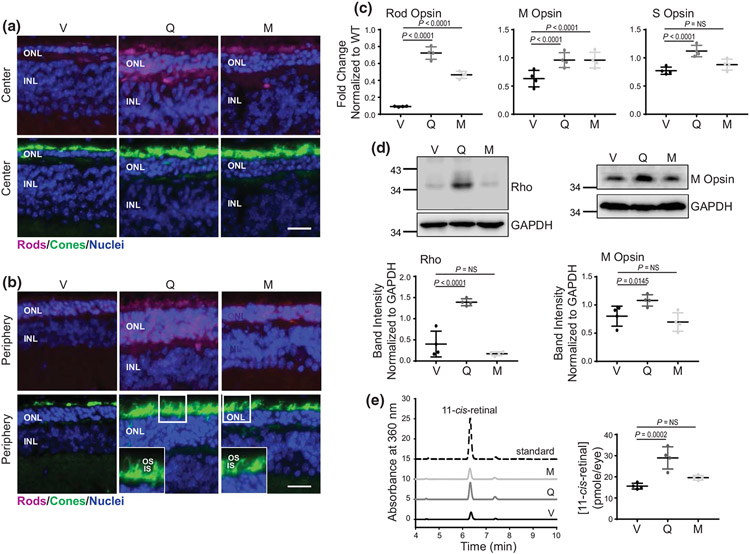
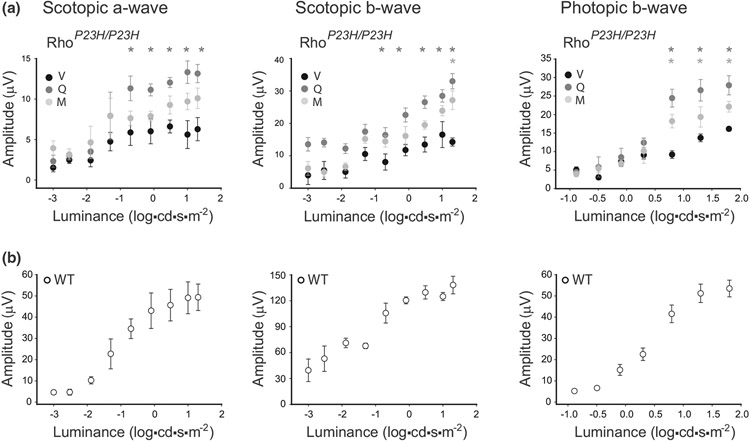
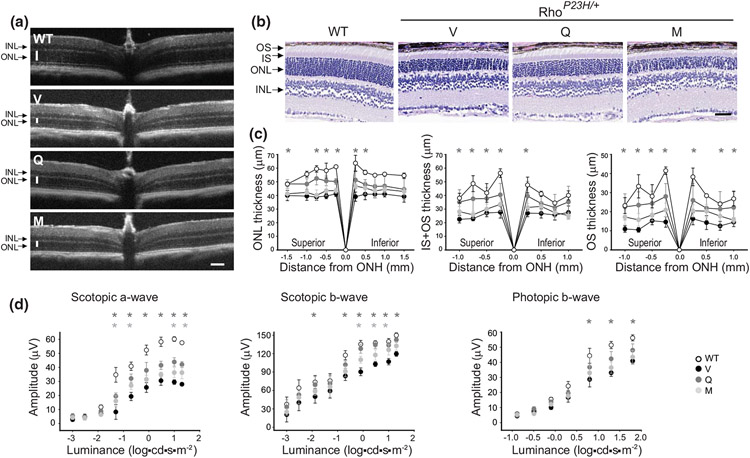
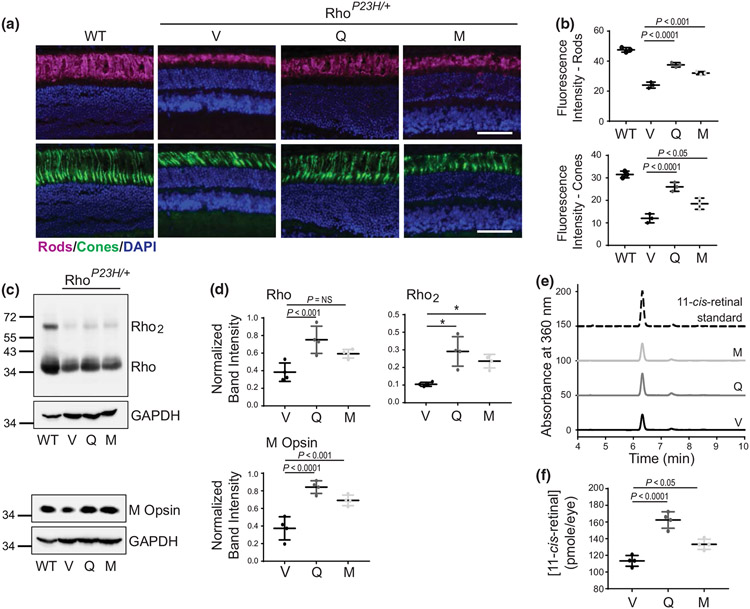
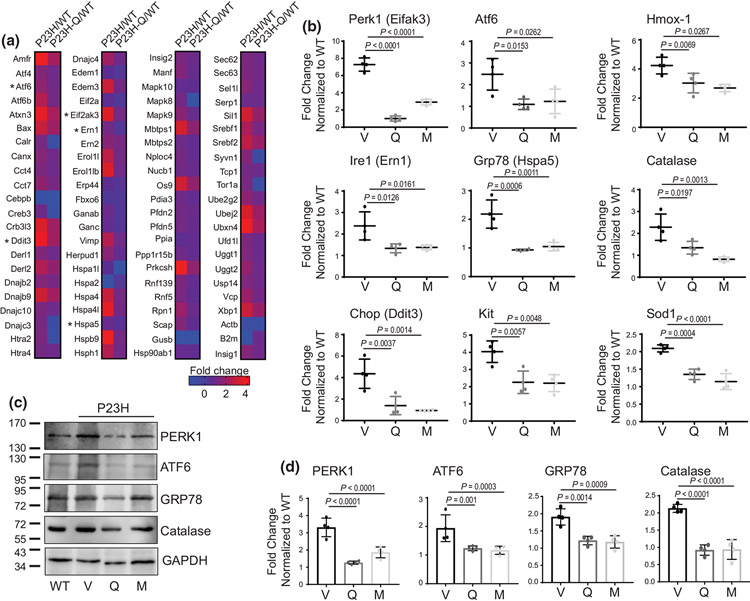
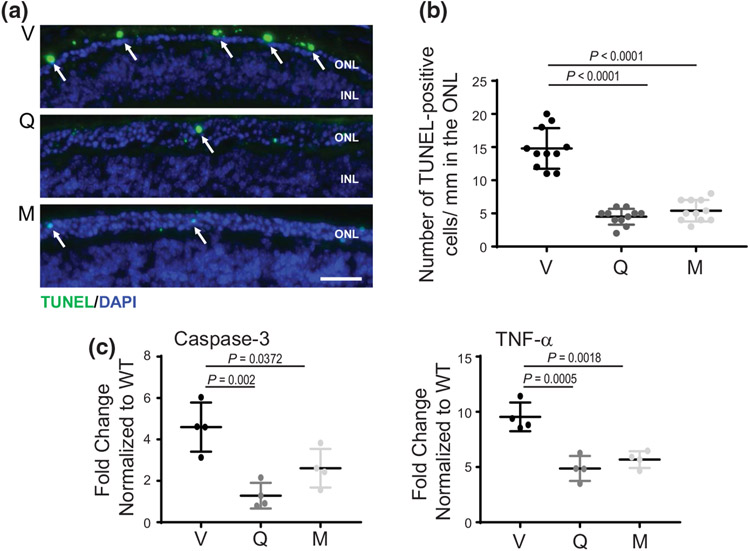
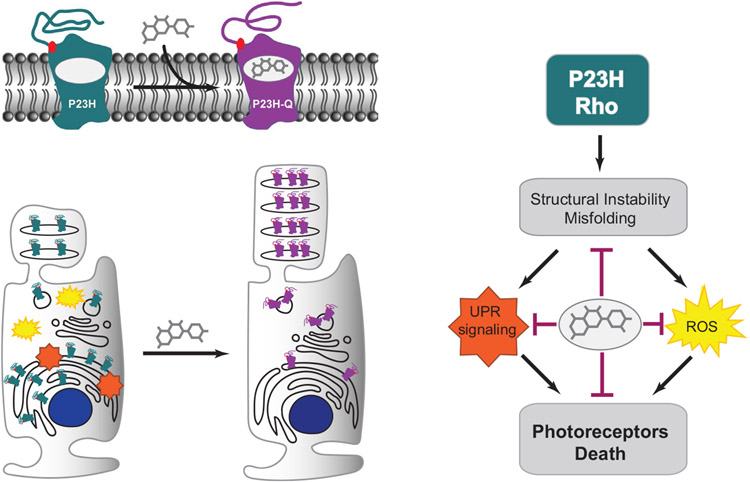
The balanced homeostasis of the G protein-coupled receptor (GPCR), rhodopsin (Rho), is required for vision. Misfolding mutations in Rho cause photoreceptor death, leading to retinitis pigmentosa (RP) and consequently blindness. With no cure currently available, the development of efficient therapy for RP is an urgent need. Pharmacological supplementation with molecular chaperones, including flavonoids, improves stability, folding, and membrane targeting of the RP Rho mutants in vitro. Thus, we hypothesized that flavonoids by binding to P23H Rho and enhancing its conformational stability could mitigate detrimental effects of this mutation on retinal health. In this work, we evaluated the pharmacological potential of two model flavonoids, quercetin and myricetin, by using in silico, in vitro, and in vivo models of P23H Rho. Our computational analysis showed that quercetin could interact within the orthosteric binding pocket of P23H Rho and shift the conformation of its N-terminal loop toward the wild type (WT)-like state. Quercetin added to the NIH-3T3 cells stably expressing P23H Rho increased the stability of this receptor and improved its function. Systemic administration of quercetin to P23H Rho knock-in mice substantially improved retinal morphology and function, which was associated with an increase in levels of Rho and cone opsins. In addition, treatment with quercetin resulted in downregulation of the UPR signaling and oxidative stress-related markers. This study unravels the pharmacological potential of quercetin to slow down the progression of photoreceptor death in Rho-related RP and highlights its prospective as a lead compound to develop a novel therapeutic remedy to counter RP pathology.
Keywords: cone opsin; flavonoid; photoreceptor; retinal degeneration; retinitis pigmentosa; rhodopsin.
© 2022 Wiley Periodicals LLC.
Conflict of interest statement
CONFLICT OF INTEREST
The authors declare that they have no conflict of interest with the contents of this article.
Figures
Similar articles
-
ATF6 is required for efficient rhodopsin clearance and retinal homeostasis in the P23H rho retinitis pigmentosa mouse model.Sci Rep. 2021 Aug 11;11(1):16356. doi: 10.1038/s41598-021-95895-7. Sci Rep. 2021. PMID: 34381136 Free PMC article.
-
The heat-shock response co-inducer arimoclomol protects against retinal degeneration in rhodopsin retinitis pigmentosa.Cell Death Dis. 2014 May 22;5(5):e1236. doi: 10.1038/cddis.2014.214. Cell Death Dis. 2014. PMID: 24853414 Free PMC article.
-
Flavonoids enhance rod opsin stability, folding, and self-association by directly binding to ligand-free opsin and modulating its conformation.J Biol Chem. 2019 May 17;294(20):8101-8122. doi: 10.1074/jbc.RA119.007808. Epub 2019 Apr 3. J Biol Chem. 2019. PMID: 30944172 Free PMC article.
-
Rhodopsin as a Molecular Target to Mitigate Retinitis Pigmentosa.Adv Exp Med Biol. 2022;1371:61-77. doi: 10.1007/5584_2021_682. Adv Exp Med Biol. 2022. PMID: 34962636 Review.
-
Targeting the Proteostasis Network in Rhodopsin Retinitis Pigmentosa.Adv Exp Med Biol. 2016;854:479-84. doi: 10.1007/978-3-319-17121-0_64. Adv Exp Med Biol. 2016. PMID: 26427449 Free PMC article. Review.
Cited by
-
Rhodopsin-associated retinal dystrophy: Disease mechanisms and therapeutic strategies.Front Neurosci. 2023 Apr 3;17:1132179. doi: 10.3389/fnins.2023.1132179. eCollection 2023. Front Neurosci. 2023. PMID: 37077319 Free PMC article. Review.
-
Molecular mechanisms underlying inherited photoreceptor degeneration as targets for therapeutic intervention.Front Cell Neurosci. 2024 Feb 2;18:1343544. doi: 10.3389/fncel.2024.1343544. eCollection 2024. Front Cell Neurosci. 2024. PMID: 38370034 Free PMC article. Review.
-
Chromenone derivatives as novel pharmacological chaperones for retinitis pigmentosa-linked rod opsin mutants.Hum Mol Genet. 2022 Oct 10;31(20):3439-3457. doi: 10.1093/hmg/ddac125. Hum Mol Genet. 2022. PMID: 35642742 Free PMC article.
-
Mechanisms of Rhodopsin-Related Inherited Retinal Degeneration and Pharmacological Treatment Strategies.Cells. 2025 Jan 4;14(1):49. doi: 10.3390/cells14010049. Cells. 2025. PMID: 39791750 Free PMC article. Review.
-
Mutant protein chemical rescue: From mechanisms to therapeutics.J Biol Chem. 2025 Apr;301(4):108417. doi: 10.1016/j.jbc.2025.108417. Epub 2025 Mar 18. J Biol Chem. 2025. PMID: 40113044 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
