

Rare catastrophes and evolutionary legacies: human germline gene variants in MLKL and the necroptosis signalling pathway
- PMID: 35166320
- PMCID: PMC9022980
- DOI: 10.1042/BST20210517
Rare catastrophes and evolutionary legacies: human germline gene variants in MLKL and the necroptosis signalling pathway
Abstract
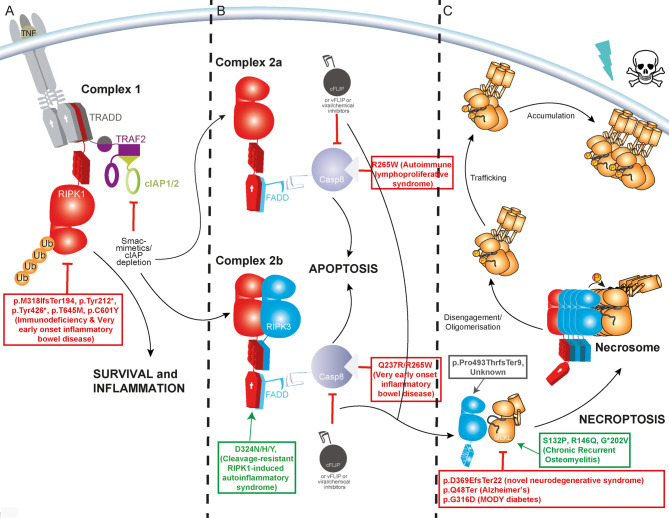
Programmed cell death has long been characterised as a key player in the development of human disease. Necroptosis is a lytic form of programmed cell death that is universally mediated by the effector protein mixed lineage kinase domain-like (MLKL), a pseudokinase. MLKL's activating kinase, receptor interacting protein kinase 3 (RIPK3), is itself activated within context specific scaffolds of receptor interacting protein kinase 1 (RIPK1), Z-DNA Binding Protein-1 (ZBP1) or TIR domain-containing adaptor inducing interferon-β (TRIF). These core necroptosis modulating proteins have been comprehensively revealed as potent drivers and suppressors of disease in inbred mouse strains. However, their roles in human disease within the 'real world' of diverse genetic backgrounds, natural infection and environmental challenges remains less well understood. Over 20 unique disease-associated human germline gene variants in this core necroptotic machinery have been reported in the literature and human clinico-genetics databases like ClinVar to date. In this review, we provide an overview of these human gene variants, with an emphasis on those encoding MLKL. These experiments of nature have the potential to not only enrich our understanding of the basic biology of necroptosis, but offer important population level insights into which clinical indications stand to benefit most from necroptosis-targeted drugs.
Keywords: MLKL; loss of function gene variant; missense gene variant; necroptosis; pathogenic mutation.
© 2022 The Author(s).
Conflict of interest statement
S.E.G. and J.M.H. contribute to a project developing necroptosis inhibitors in collaboration with Anaxis Pty Ltd.
Figures

Similar articles
-
Cell-type dependence of necroptosis pathways triggered by viral infection.FEBS J. 2024 Jun;291(11):2388-2404. doi: 10.1111/febs.17045. Epub 2024 Jan 9. FEBS J. 2024. PMID: 38145501
-
Z-DNA/RNA Binding Protein 1 Senses Mitochondrial DNA to Induce Receptor-Interacting Protein Kinase-3/Mixed Lineage Kinase Domain-Like-Driven Necroptosis in Developmental Sevoflurane Neurotoxicity.Neuroscience. 2022 Dec 15;507:99-111. doi: 10.1016/j.neuroscience.2022.11.005. Epub 2022 Nov 10. Neuroscience. 2022. PMID: 36370933
-
Influenza Virus Z-RNAs Induce ZBP1-Mediated Necroptosis.Cell. 2020 Mar 19;180(6):1115-1129.e13. doi: 10.1016/j.cell.2020.02.050. Cell. 2020. PMID: 32200799 Free PMC article.
-
The Role of the Key Effector of Necroptotic Cell Death, MLKL, in Mouse Models of Disease.Biomolecules. 2021 May 28;11(6):803. doi: 10.3390/biom11060803. Biomolecules. 2021. PMID: 34071602 Free PMC article. Review.
-
Protein shapeshifting in necroptotic cell death signaling.Trends Biochem Sci. 2025 Feb;50(2):92-105. doi: 10.1016/j.tibs.2024.11.006. Epub 2024 Dec 26. Trends Biochem Sci. 2025. PMID: 39730228 Review.
Cited by
-
Mediators of necroptosis: from cell death to metabolic regulation.EMBO Mol Med. 2024 Feb;16(2):219-237. doi: 10.1038/s44321-023-00011-z. Epub 2024 Jan 9. EMBO Mol Med. 2024. PMID: 38195700 Free PMC article. Review.
-
Inhibitors identify an auxiliary role for mTOR signalling in necroptosis execution downstream of MLKL activation.Biochem J. 2024 Sep 4;481(17):1125-1142. doi: 10.1042/BCJ20240255. Biochem J. 2024. PMID: 39136677 Free PMC article.
-
A common human MLKL polymorphism confers resistance to negative regulation by phosphorylation.Nat Commun. 2023 Sep 28;14(1):6046. doi: 10.1038/s41467-023-41724-6. Nat Commun. 2023. PMID: 37770424 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous