

The maize single-nucleus transcriptome comprehensively describes signaling networks governing movement and development of grass stomata
- PMID: 35166333
- PMCID: PMC9048877
- DOI: 10.1093/plcell/koac047
The maize single-nucleus transcriptome comprehensively describes signaling networks governing movement and development of grass stomata
Abstract
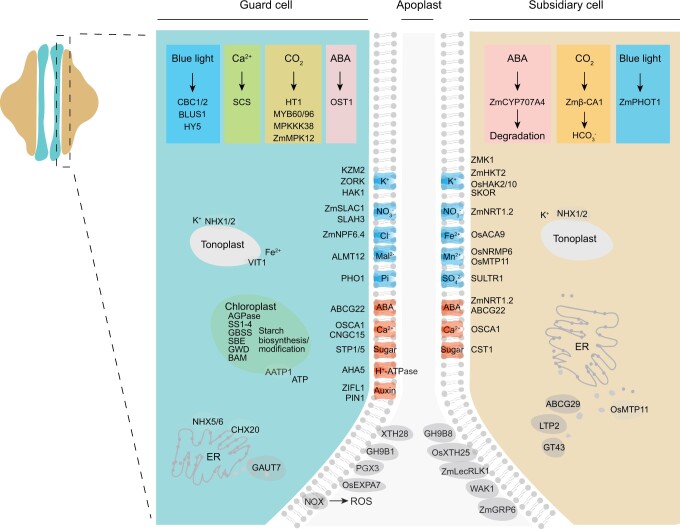
The unique morphology of grass stomata enables rapid responses to environmental changes. Deciphering the basis for these responses is critical for improving food security. We have developed a planta platform of single-nucleus RNA-sequencing by combined fluorescence-activated nuclei flow sorting, and used it to identify cell types in mature and developing stomata from 33,098 nuclei of the maize epidermis-enriched tissues. Guard cells (GCs) and subsidiary cells (SCs) displayed differential expression of genes, besides those encoding transporters, involved in the abscisic acid, CO2, Ca2+, starch metabolism, and blue light signaling pathways, implicating coordinated signal integration in speedy stomatal responses, and of genes affecting cell wall plasticity, implying a more sophisticated relationship between GCs and SCs in stomatal development and dumbbell-shaped guard cell formation. The trajectory of stomatal development identified in young tissues, and by comparison to the bulk RNA-seq data of the MUTE defective mutant in stomatal development, confirmed known features, and shed light on key participants in stomatal development. Our study provides a valuable, comprehensive, and fundamental foundation for further insights into grass stomatal function.
© American Society of Plant Biologists 2022. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures





Comment in
-
Single-nucleus transcriptomics for an integrative view of grass stomatal processes.Plant Cell. 2022 Apr 26;34(5):1882-1883. doi: 10.1093/plcell/koac054. Plant Cell. 2022. PMID: 35288755 Free PMC article. No abstract available.
References
-
- Acevedo-Garcia J, Kusch S, Panstruga R (2014) Magical mystery tour: MLO proteins in plant immunity and beyond. New Phytol 204: 273–281 - PubMed
-
- Alejandro S, Lee Y, Tohge T, Sudre D, Osorio S, Park J, Bovet L, Lee Y, Geldner N, Fernie AR (2012) AtABCG29 is a monolignol transporter involved in lignin biosynthesis. Curr Biol 22: 1207–1212 - PubMed
-
- Atmodjo MA, Sakuragi Y, Zhu X, Burrell AJ, Mohanty SS, Atwood JA 3rd, Orlando R, Scheller HV, Mohnen D (2011) Galacturonosyltransferase (GAUT)1 and GAUT7 are the core of a plant cell wall pectin biosynthetic homogalacturonan:galacturonosyltransferase complex. Proc Natl Acad Sci USA 108: 20225–20230 - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
