

Structural effects of m6A modification of the Xist A-repeat AUCG tetraloop and its recognition by YTHDC1
- PMID: 35166835
- PMCID: PMC8887474
- DOI: 10.1093/nar/gkac080
Structural effects of m6A modification of the Xist A-repeat AUCG tetraloop and its recognition by YTHDC1
Abstract
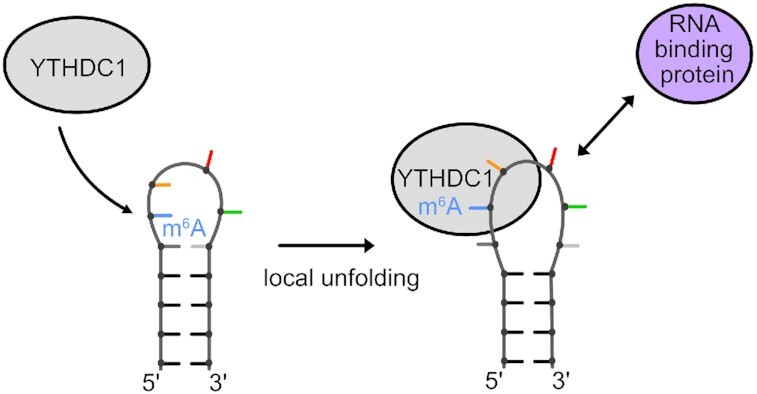
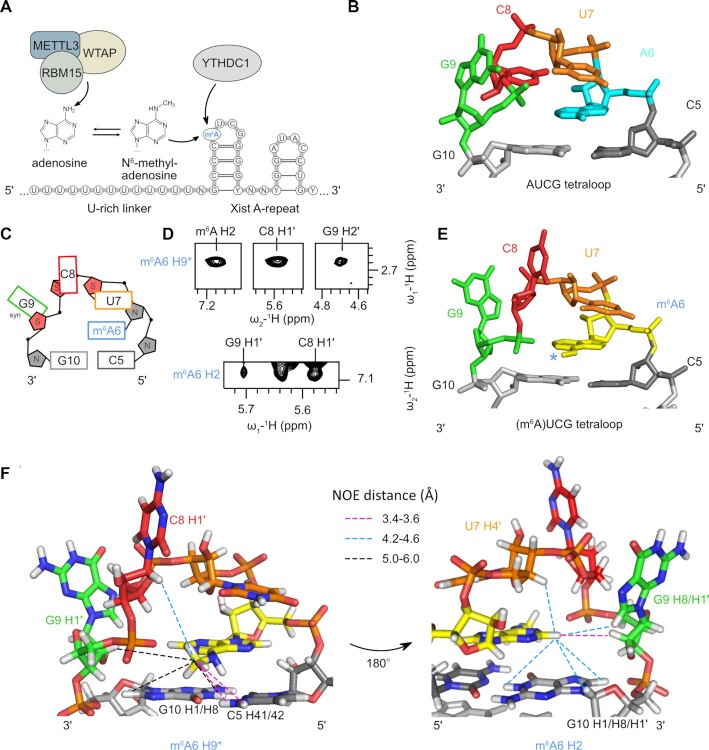
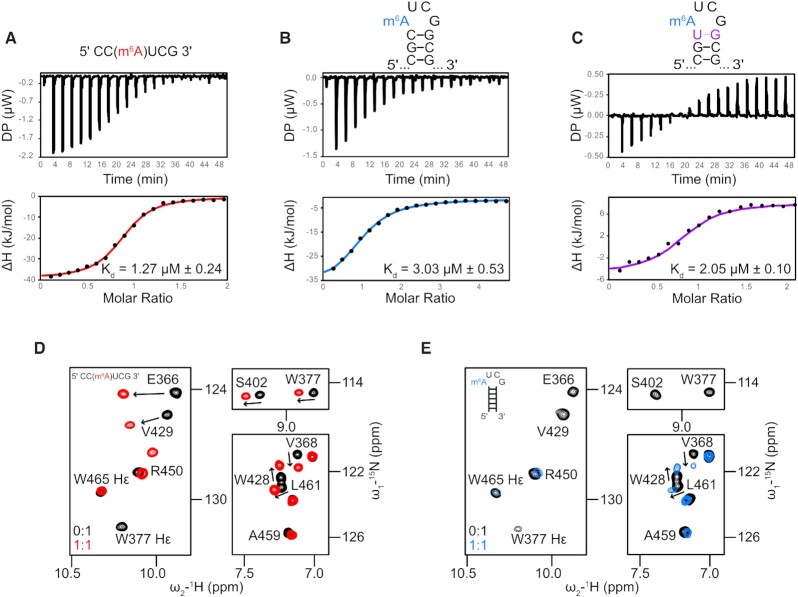
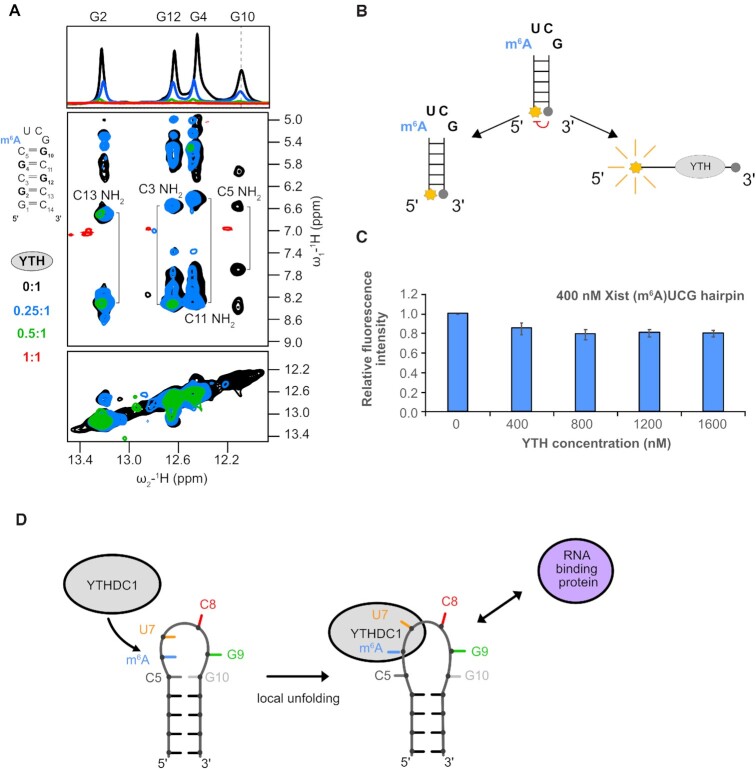
The A-repeat region of the lncRNA Xist is critical for X inactivation and harbors several N6-methyladenosine (m6A) modifications. How the m6A modification affects the conformation of the conserved AUCG tetraloop hairpin of the A-repeats and how it can be recognized by the YTHDC1 reader protein is unknown. Here, we report the NMR solution structure of the (m6A)UCG hairpin, which reveals that the m6A base extends 5' stacking of the A-form helical stem, resembling the unmethylated AUCG tetraloop. A crystal structure of YTHDC1 bound to the (m6A)UCG tetraloop shows that the (m6A)UC nucleotides are recognized by the YTH domain of YTHDC1 in a single-stranded conformation. The m6A base inserts into the aromatic cage and the U and C bases interact with a flanking charged surface region, resembling the recognition of single-stranded m6A RNA ligands. Notably, NMR and fluorescence quenching experiments show that the binding requires local unfolding of the upper stem region of the (m6A)UCG hairpin. Our data show that m6A can be readily accommodated in hairpin loop regions, but recognition by YTH readers requires local unfolding of flanking stem regions. This suggests how m6A modifications may regulate lncRNA function by modulating RNA structure.
© The Author(s) 2022. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures

Similar articles
-
Systematic molecular profiling of non-native N6-substitution effects on m6A binding to the YTH domains of human RNA m6A readers in diabetes.Biophys Chem. 2025 May-Jun;320-321:107417. doi: 10.1016/j.bpc.2025.107417. Epub 2025 Feb 18. Biophys Chem. 2025. PMID: 39987708
-
Structural basis for selective binding of m6A RNA by the YTHDC1 YTH domain.Nat Chem Biol. 2014 Nov;10(11):927-9. doi: 10.1038/nchembio.1654. Epub 2014 Sep 21. Nat Chem Biol. 2014. PMID: 25242552
-
m(6)A: Signaling for mRNA splicing.RNA Biol. 2016 Sep;13(9):756-9. doi: 10.1080/15476286.2016.1201628. Epub 2016 Jun 28. RNA Biol. 2016. PMID: 27351695 Free PMC article. Review.
-
Adenine Methylation Enhances the Conformational Flexibility of an RNA Hairpin Tetraloop.J Phys Chem B. 2024 Apr 4;128(13):3157-3166. doi: 10.1021/acs.jpcb.4c00522. Epub 2024 Mar 27. J Phys Chem B. 2024. PMID: 38535997 Free PMC article.
-
The potential role of RNA N6-methyladenosine in Cancer progression.Mol Cancer. 2020 May 12;19(1):88. doi: 10.1186/s12943-020-01204-7. Mol Cancer. 2020. PMID: 32398132 Free PMC article. Review.
Cited by
-
Long noncoding RNA MEG3 inhibits oral squamous cell carcinoma progression via GATA3.FEBS Open Bio. 2023 Jan;13(1):195-208. doi: 10.1002/2211-5463.13532. Epub 2022 Dec 18. FEBS Open Bio. 2023. PMID: 36468944 Free PMC article.
-
METTL3 drives telomere targeting of TERRA lncRNA through m6A-dependent R-loop formation: a therapeutic target for ALT-positive neuroblastoma.Nucleic Acids Res. 2024 Mar 21;52(5):2648-2671. doi: 10.1093/nar/gkad1242. Nucleic Acids Res. 2024. PMID: 38180812 Free PMC article.
-
The roles of N6-methyladenosine and its target regulatory noncoding RNAs in tumors: classification, mechanisms, and potential therapeutic implications.Exp Mol Med. 2023 Mar;55(3):487-501. doi: 10.1038/s12276-023-00944-y. Epub 2023 Mar 1. Exp Mol Med. 2023. PMID: 36854773 Free PMC article. Review.
-
The chromatin-associated RNAs in gene regulation and cancer.Mol Cancer. 2023 Feb 7;22(1):27. doi: 10.1186/s12943-023-01724-y. Mol Cancer. 2023. PMID: 36750826 Free PMC article. Review.
-
RNA: The Unsuspected Conductor in the Orchestra of Macromolecular Crowding.Chem Rev. 2024 Apr 24;124(8):4734-4777. doi: 10.1021/acs.chemrev.3c00575. Epub 2024 Apr 5. Chem Rev. 2024. PMID: 38579177 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
