

Butterfly eyespots evolved via cooption of an ancestral gene-regulatory network that also patterns antennae, legs, and wings
- PMID: 35169073
- PMCID: PMC8872758
- DOI: 10.1073/pnas.2108661119
Butterfly eyespots evolved via cooption of an ancestral gene-regulatory network that also patterns antennae, legs, and wings
Abstract
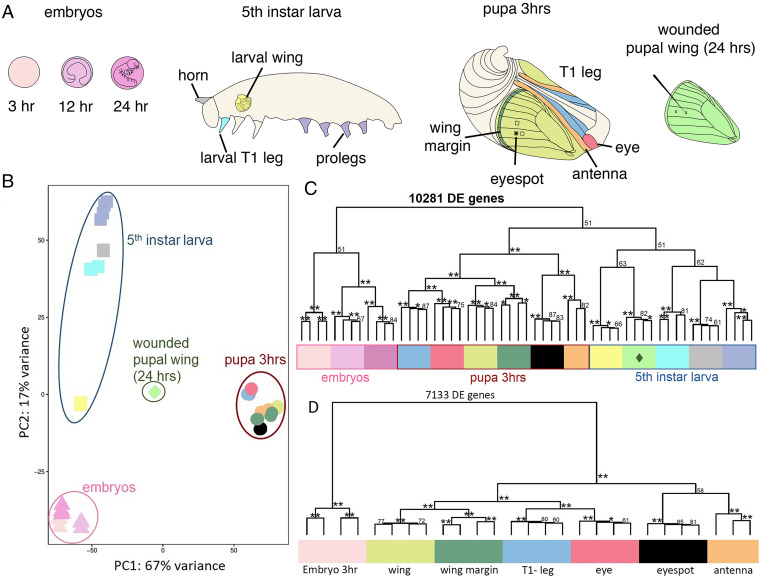
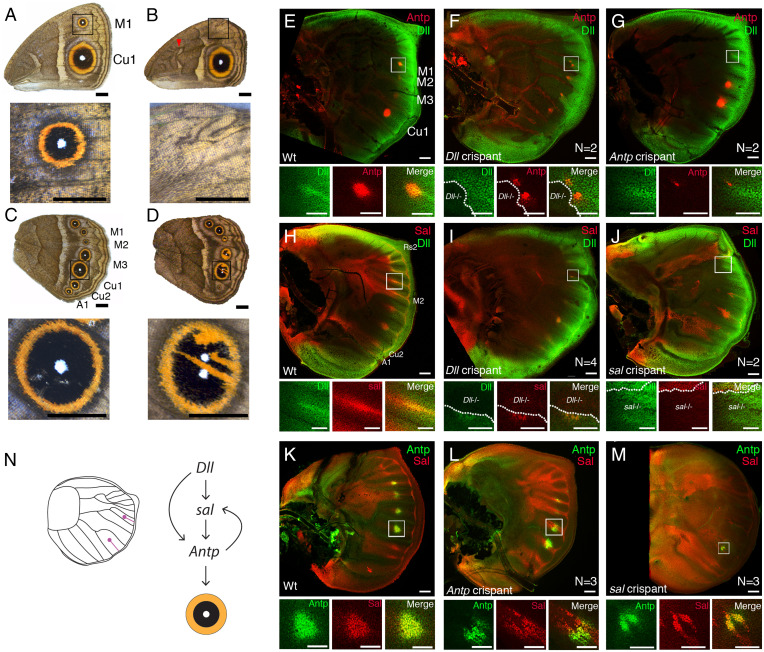
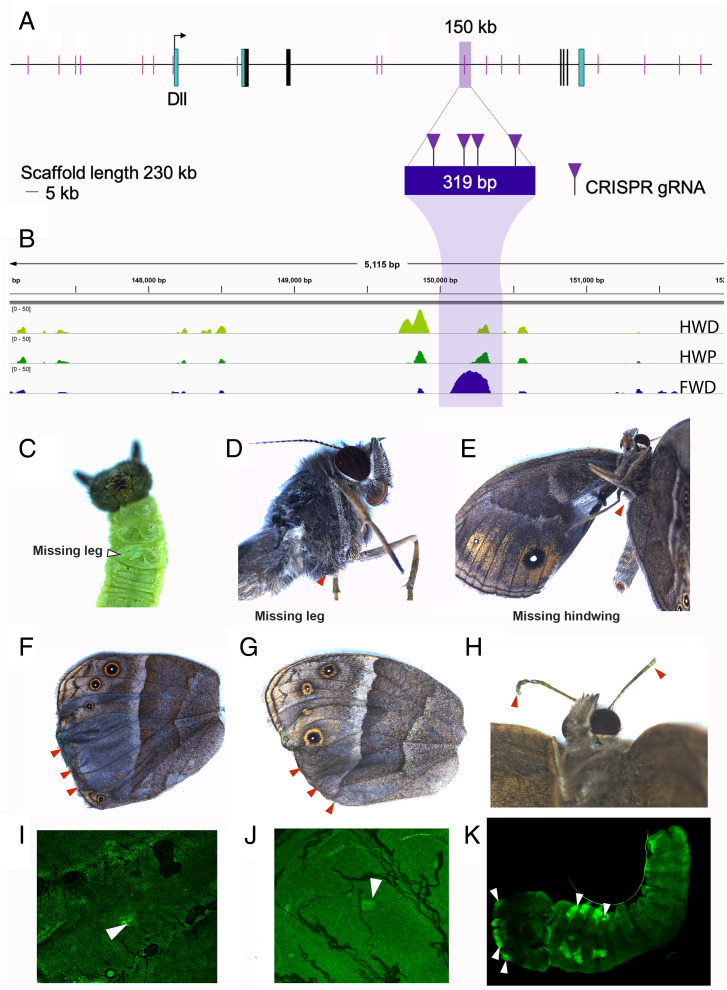
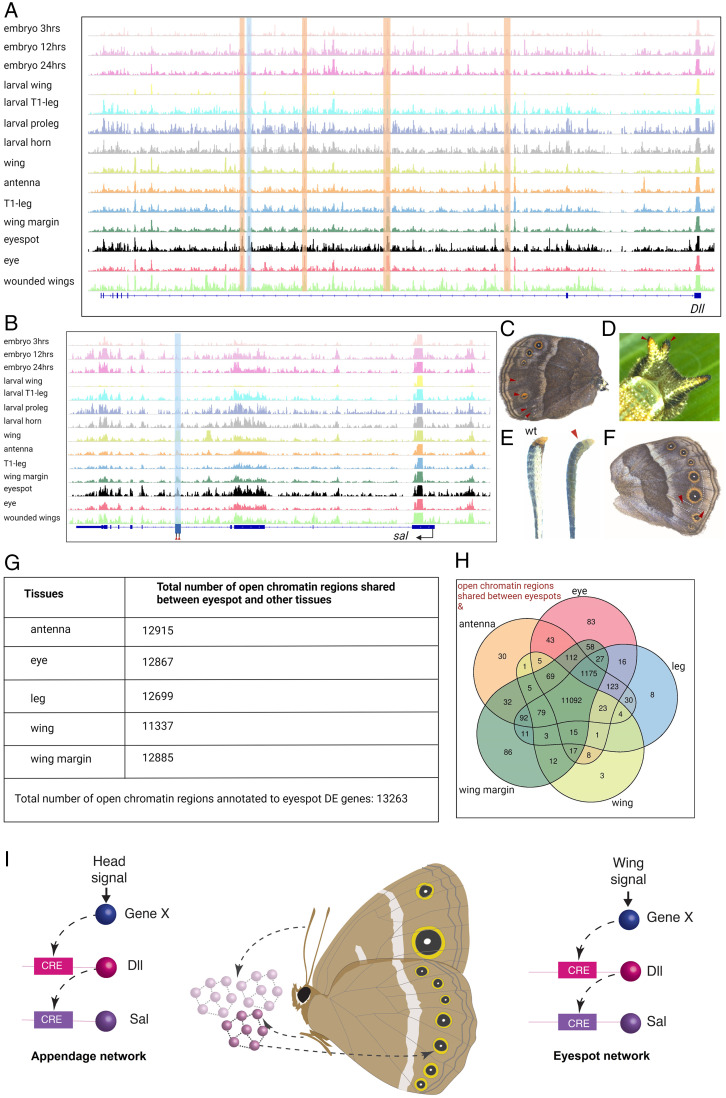
Butterfly eyespots are beautiful novel traits with an unknown developmental origin. Here we show that eyespots likely originated via cooption of parts of an ancestral appendage gene-regulatory network (GRN) to novel locations on the wing. Using comparative transcriptome analysis, we show that eyespots cluster most closely with antennae, relative to multiple other tissues. Furthermore, three genes essential for eyespot development, Distal-less (Dll), spalt (sal), and Antennapedia (Antp), share similar regulatory connections as those observed in the antennal GRN. CRISPR knockout of cis-regulatory elements (CREs) for Dll and sal led to the loss of eyespots, antennae, legs, and also wings, demonstrating that these CREs are highly pleiotropic. We conclude that eyespots likely reused an ancient GRN for their development, a network also previously implicated in the development of antennae, legs, and wings.
Keywords: evolutionary biology; evolutionary developmental biology; genetics.
Copyright © 2022 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Carroll S. B., et al. , Pattern formation and eyespot determination in butterfly wings. Science 265, 109–114 (1994). - PubMed
-
- Held L. I., Rethinking butterfly eyespots. Evol. Biol. 40, 158–168 (2013).
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
