

IRE1α drives lung epithelial progenitor dysfunction to establish a niche for pulmonary fibrosis
- PMID: 35170357
- PMCID: PMC8957349
- DOI: 10.1152/ajplung.00408.2021
IRE1α drives lung epithelial progenitor dysfunction to establish a niche for pulmonary fibrosis
Abstract
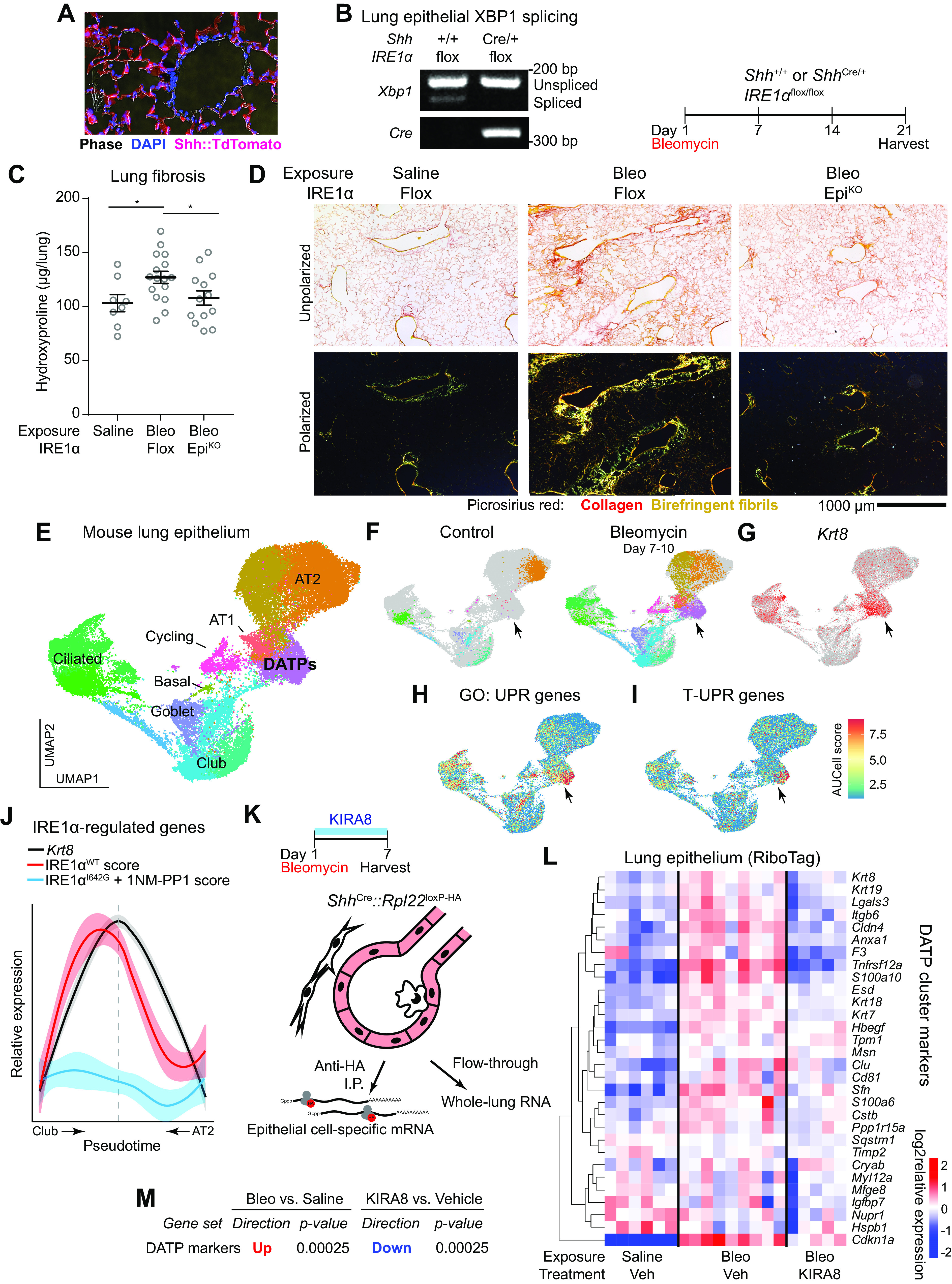
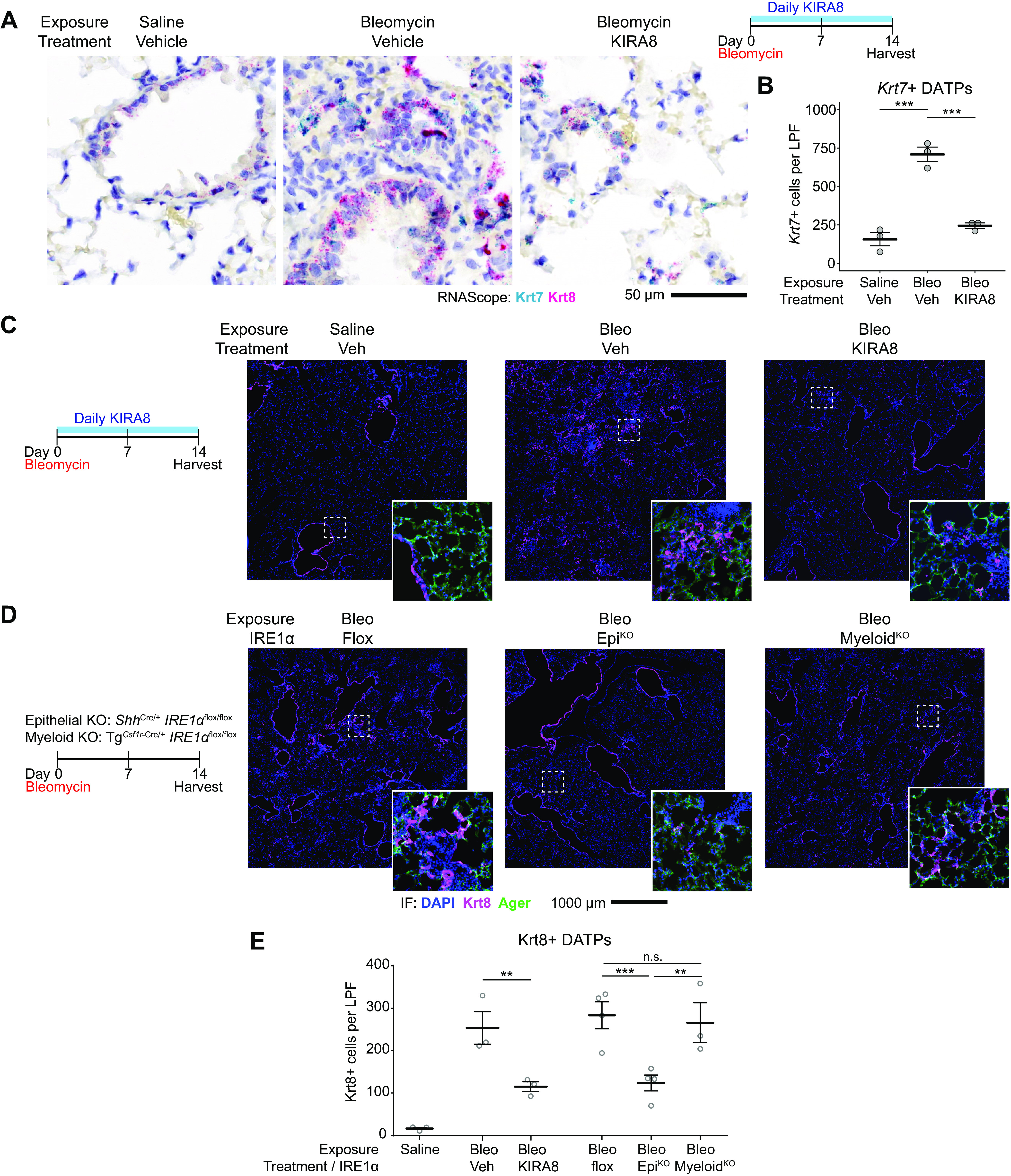
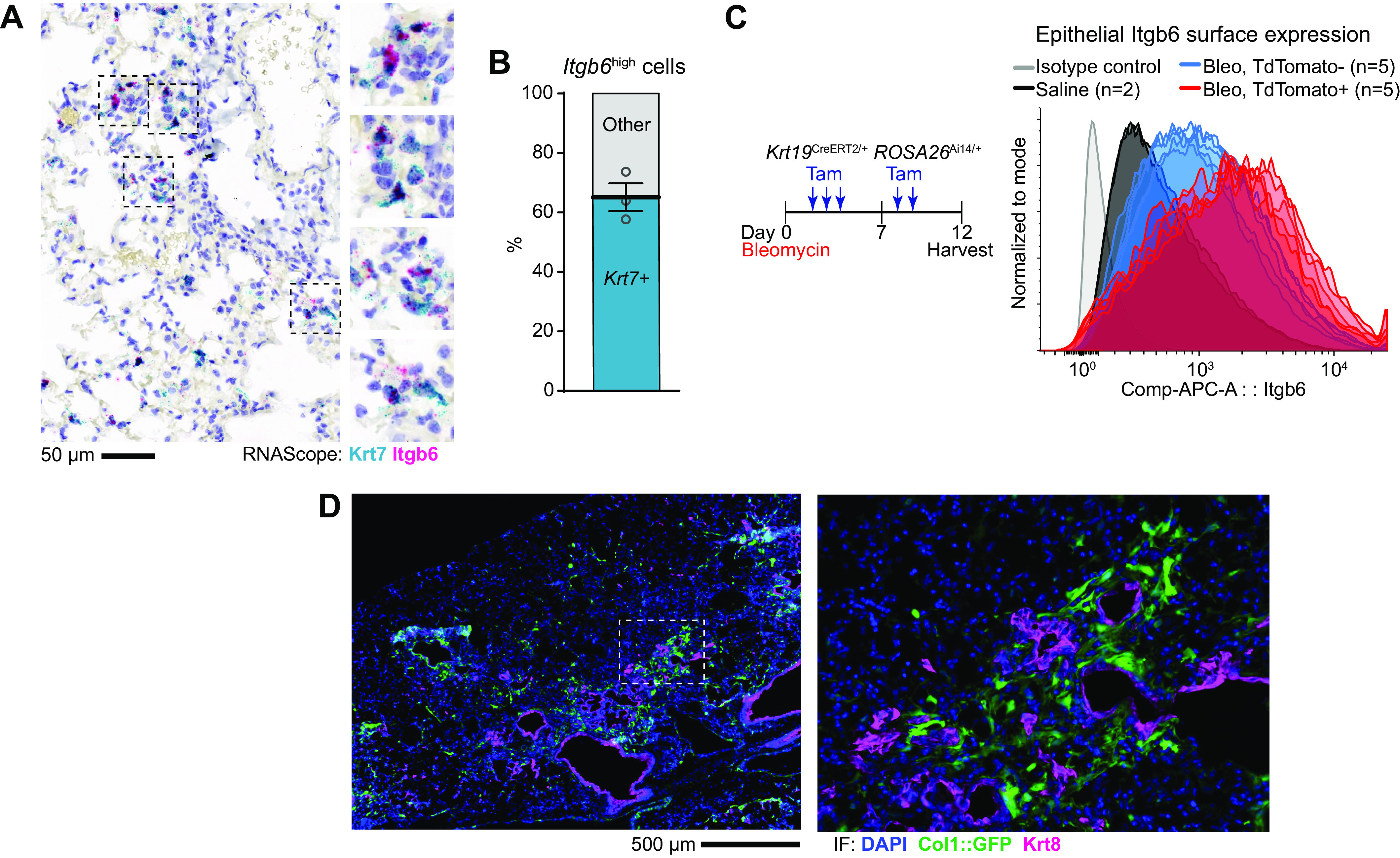
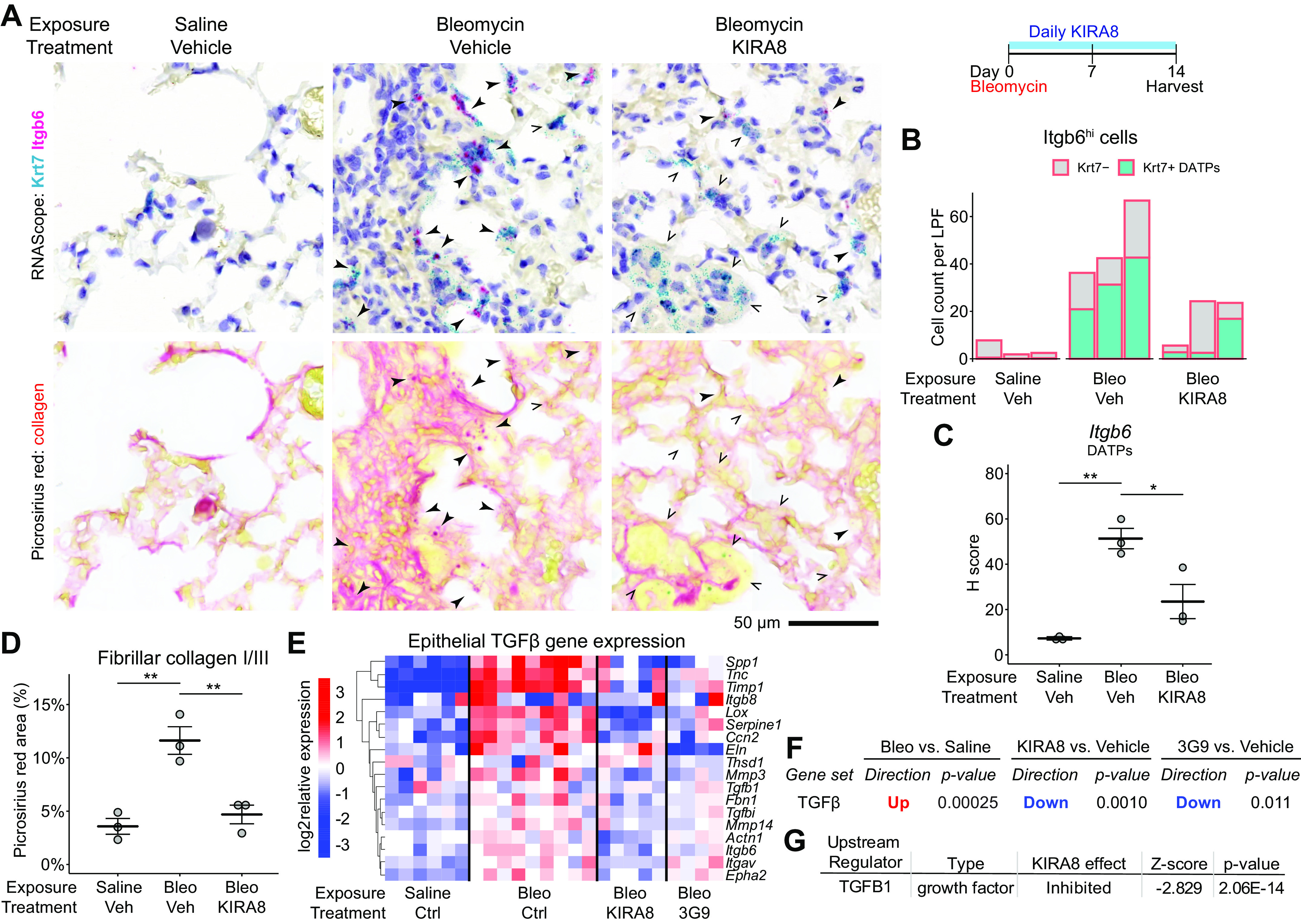
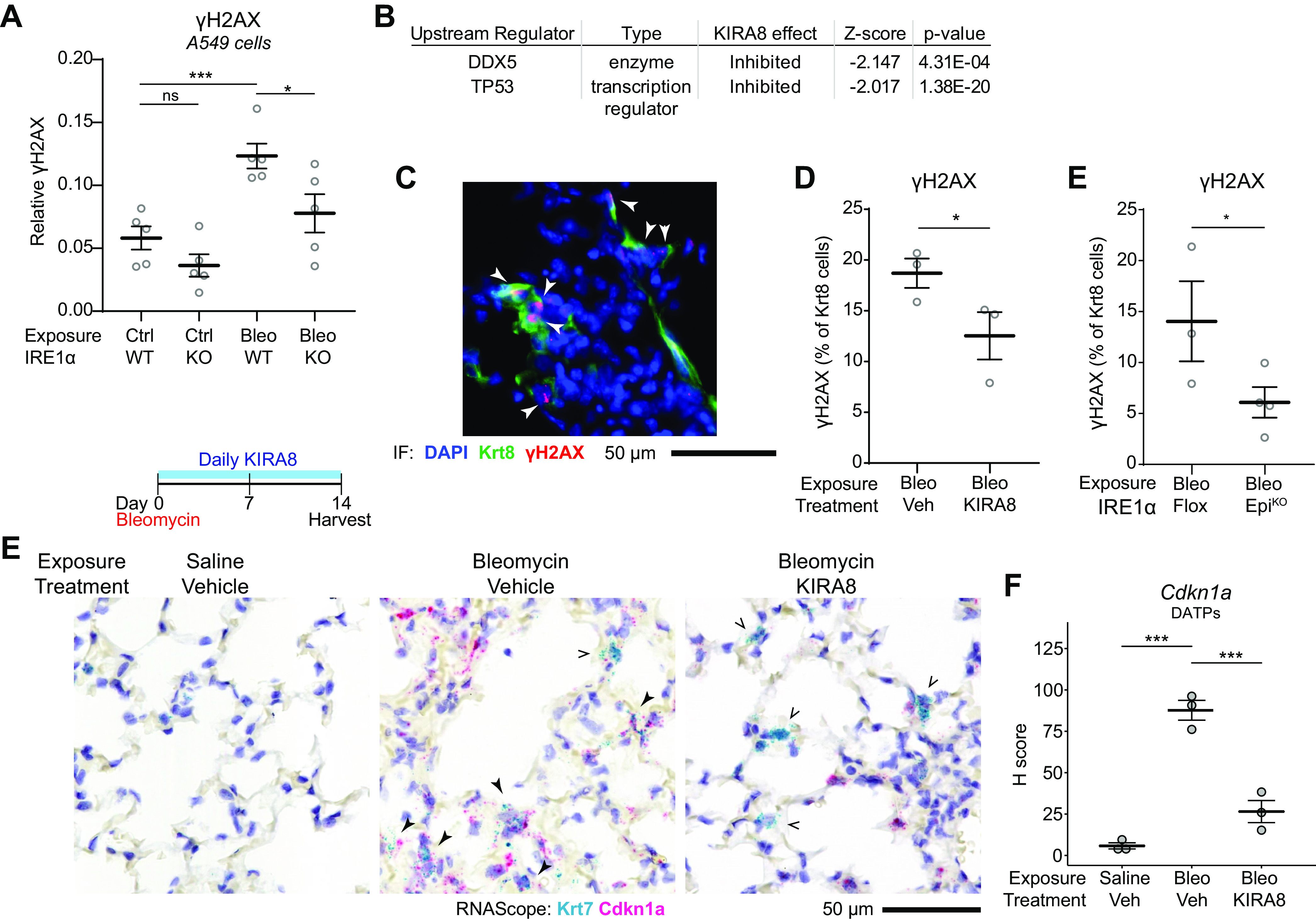
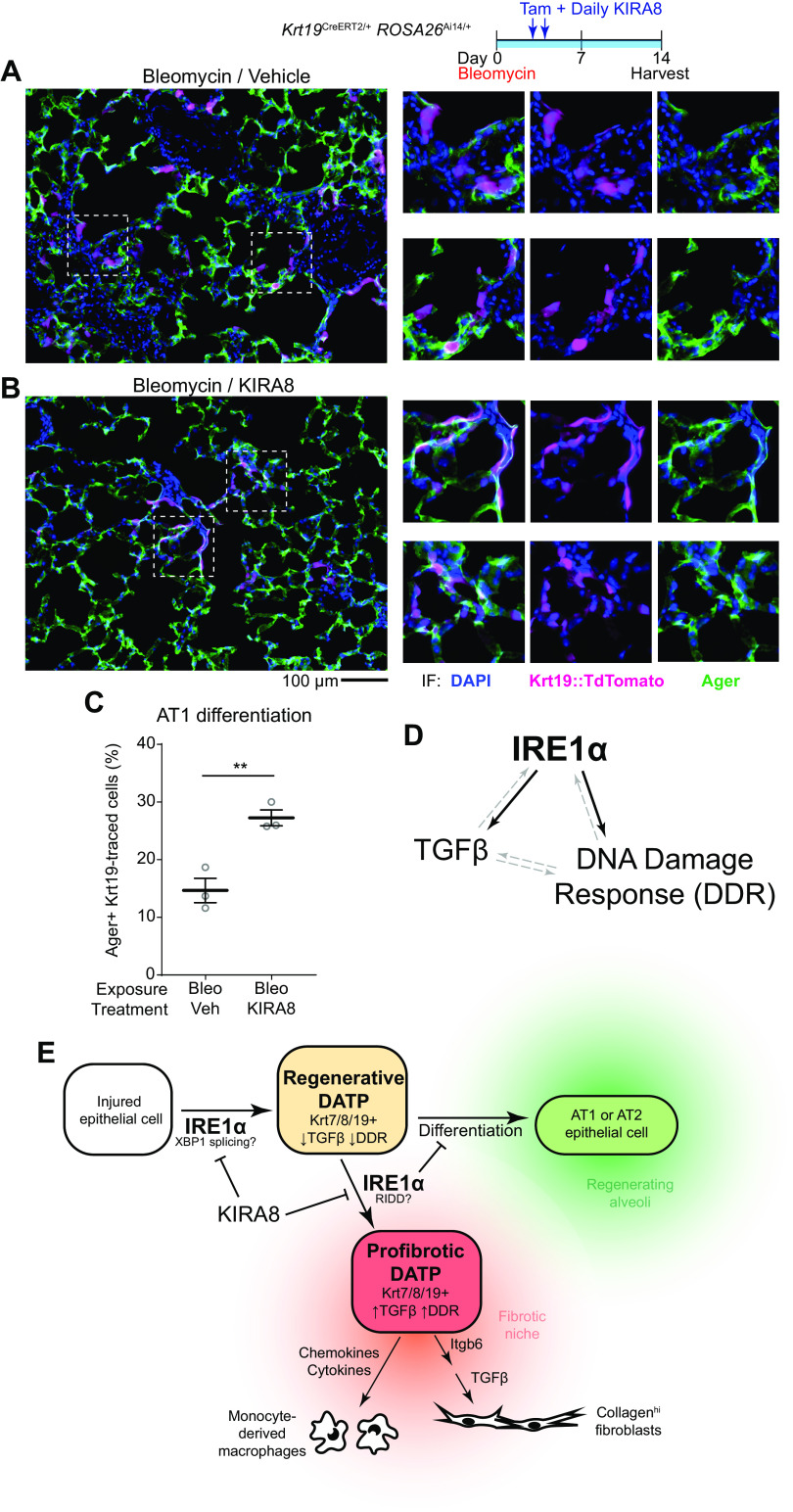
After lung injury, damage-associated transient progenitors (DATPs) emerge, representing a transitional state between injured epithelial cells and newly regenerated alveoli. DATPs express profibrotic genes, suggesting that they might promote idiopathic pulmonary fibrosis (IPF). However, the molecular pathways that induce and/or maintain DATPs are incompletely understood. Here we show that the bifunctional kinase/RNase-IRE1α-a central mediator of the unfolded protein response (UPR) to endoplasmic reticulum (ER) stress is a critical promoter of DATP abundance and function. Administration of a nanomolar-potent, monoselective kinase inhibitor of IRE1α (KIRA8)-or conditional epithelial IRE1α gene knockout-both reduce DATP cell number and fibrosis in the bleomycin model, indicating that IRE1α cell-autonomously promotes transition into the DATP state. IRE1α enhances the profibrotic phenotype of DATPs since KIRA8 decreases expression of integrin αvβ6, a key activator of transforming growth factor β (TGF-β) in pulmonary fibrosis, corresponding to decreased TGF-β-induced gene expression in the epithelium and decreased collagen accumulation around DATPs. Furthermore, IRE1α regulates DNA damage response (DDR) signaling, previously shown to promote the DATP phenotype, as IRE1α loss-of-function decreases H2AX phosphorylation, Cdkn1a (p21) expression, and DDR-associated secretory gene expression. Finally, KIRA8 treatment increases the differentiation of Krt19CreERT2-lineage-traced DATPs into type 1 alveolar epithelial cells after bleomycin injury, indicating that relief from IRE1α signaling enables DATPs to exit the transitional state. Thus, IRE1α coordinates a network of stress pathways that conspire to entrap injured cells in the DATP state. Pharmacological blockade of IRE1α signaling helps resolve the DATP state, thereby ameliorating fibrosis and promoting salutary lung regeneration.
Keywords: IRE1α; kinase inhibitor; lung regeneration; pulmonary fibrosis; unfolded protein response.
Conflict of interest statement
B.J.B. and F.R.P. are founders and equity holders of OptiKira, LLC (Cleveland, OH). No funding or chemical matter from OptiKira was used for the work described in this article. None of the other authors has any conflicts of interest, financial or otherwise, to disclose.
Figures
References
-
- Wolters PJ, Blackwell TS, Eickelberg O, Loyd JE, Kaminski N, Jenkins G, Maher TM, Molina-Molina M, Noble PW, Raghu G, Richeldi L, Schwarz MI, Selman M, Wuyts WA, Schwartz DA. Time for a change: is idiopathic pulmonary fibrosis still idiopathic and only fibrotic? Lancet Respir Med 6: 154–160, 2018. doi: 10.1016/S2213-2600(18)30007-9. - DOI - PMC - PubMed
-
- Riemondy KA, Jansing NL, Jiang P, Redente EF, Gillen AE, Fu R, Miller AJ, Spence JR, Gerber AN, Hesselberth JR, Zemans RL. Single cell RNA sequencing identifies TGFβ as a key regenerative cue following LPS-induced lung injury. JCI Insight 5: e123637, 2019. doi: 10.1172/jci.insight.123637. - DOI - PMC - PubMed
-
- Joshi N, Watanabe S, Verma R, Jablonski RP, Chen CI, Cheresh P, Markov NS, Reyfman PA, McQuattie-Pimentel AC, Sichizya L, Lu Z, Piseaux-Aillon R, Kirchenbuechler D, Flozak AS, Gottardi CJ, Cuda CM, Perlman H, Jain M, Kamp DW, Scott Budinger GR, Misharin AV. A spatially restricted fibrotic niche in pulmonary fibrosis is sustained by M-CSF/M-CSFR signalling in monocyte-derived alveolar macrophages. Eur Respir J 55: 1900646, 2020. doi: 10.1183/13993003.00646-2019. - DOI - PMC - PubMed
-
- Strunz M, Simon LM, Ansari M, Kathiriya JJ, Angelidis I, Mayr CH, Tsidiridis G, Lange M, Mattner LF, Yee M, Ogar P, Sengupta A, Kukhtevich I, Schneider R, Zhao Z, Voss C, Stoeger T, Neumann JHL, Hilgendorff A, Behr J, O’Reilly M, Lehmann M, Burgstaller G, Königshoff M, Chapman HA, Theis FJ, Schiller HB. Alveolar regeneration through a Krt8+ transitional stem cell state that persists in human lung fibrosis. Nat Commun 11: 3559, 2020. doi: 10.1038/s41467-020-17358-3. - DOI - PMC - PubMed
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous