

Astrocytic α2-Na+/K+ ATPase inhibition suppresses astrocyte reactivity and reduces neurodegeneration in a tauopathy mouse model
- PMID: 35171651
- PMCID: PMC9161722
- DOI: 10.1126/scitranslmed.abm4107
Astrocytic α2-Na+/K+ ATPase inhibition suppresses astrocyte reactivity and reduces neurodegeneration in a tauopathy mouse model
Abstract
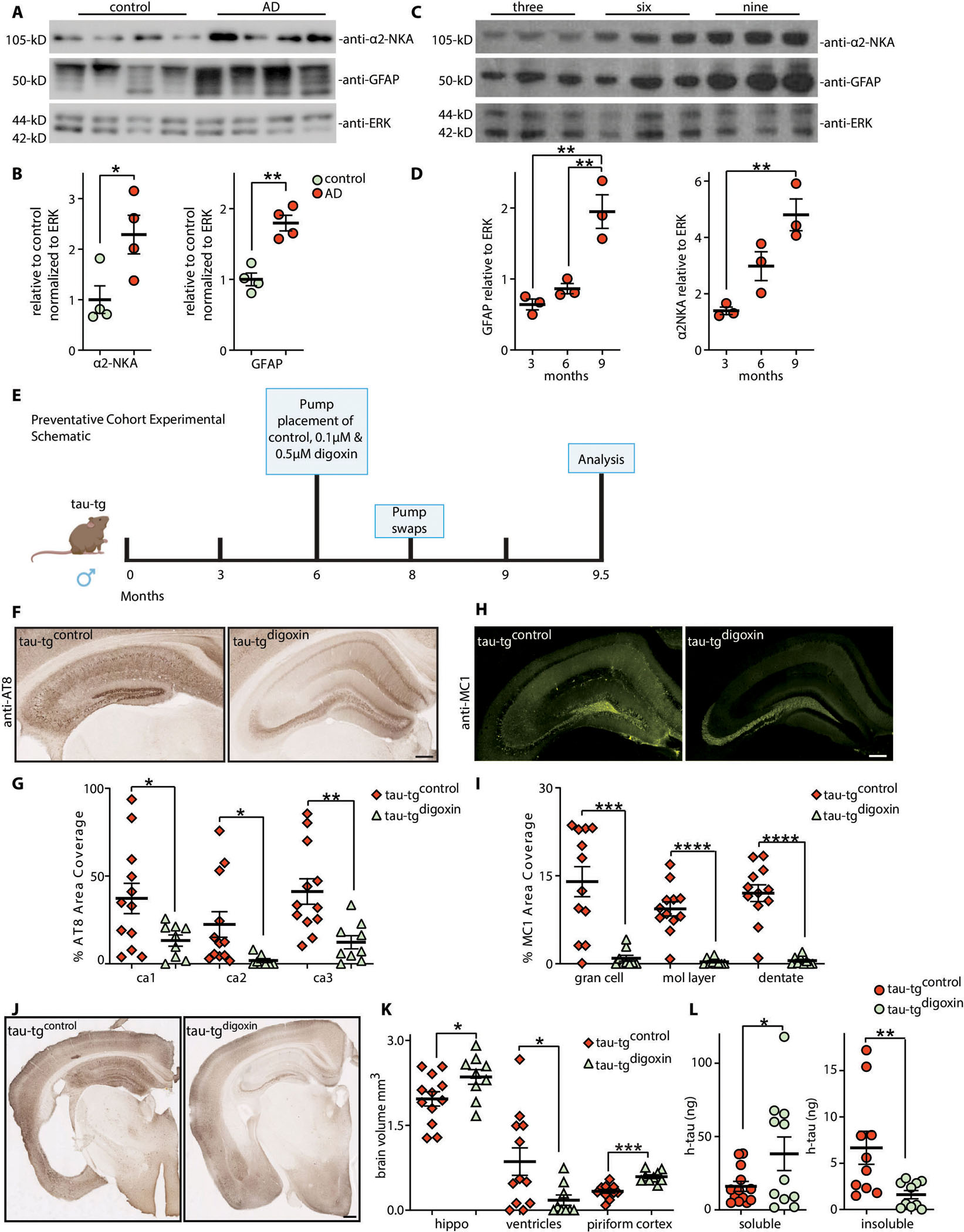
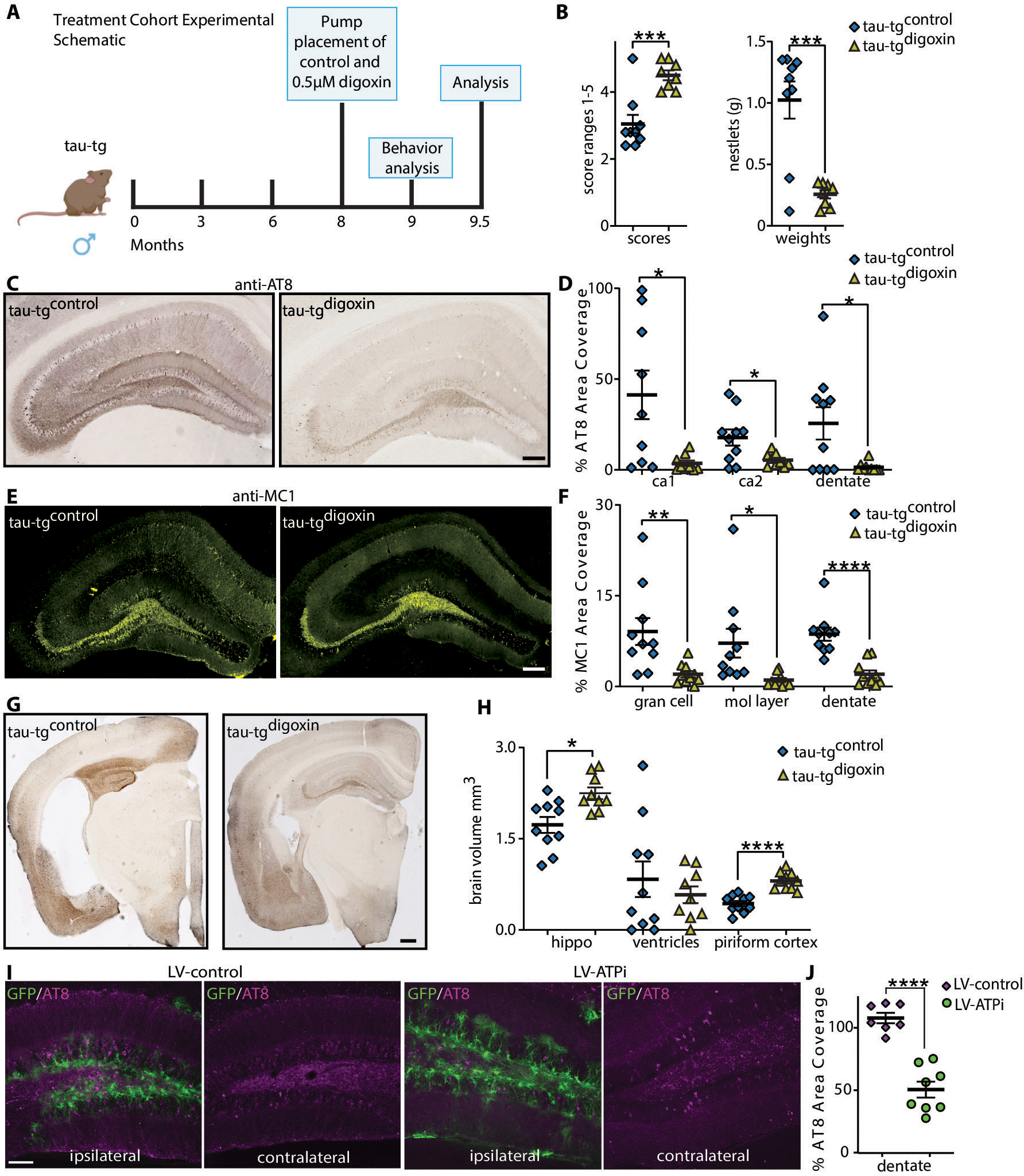
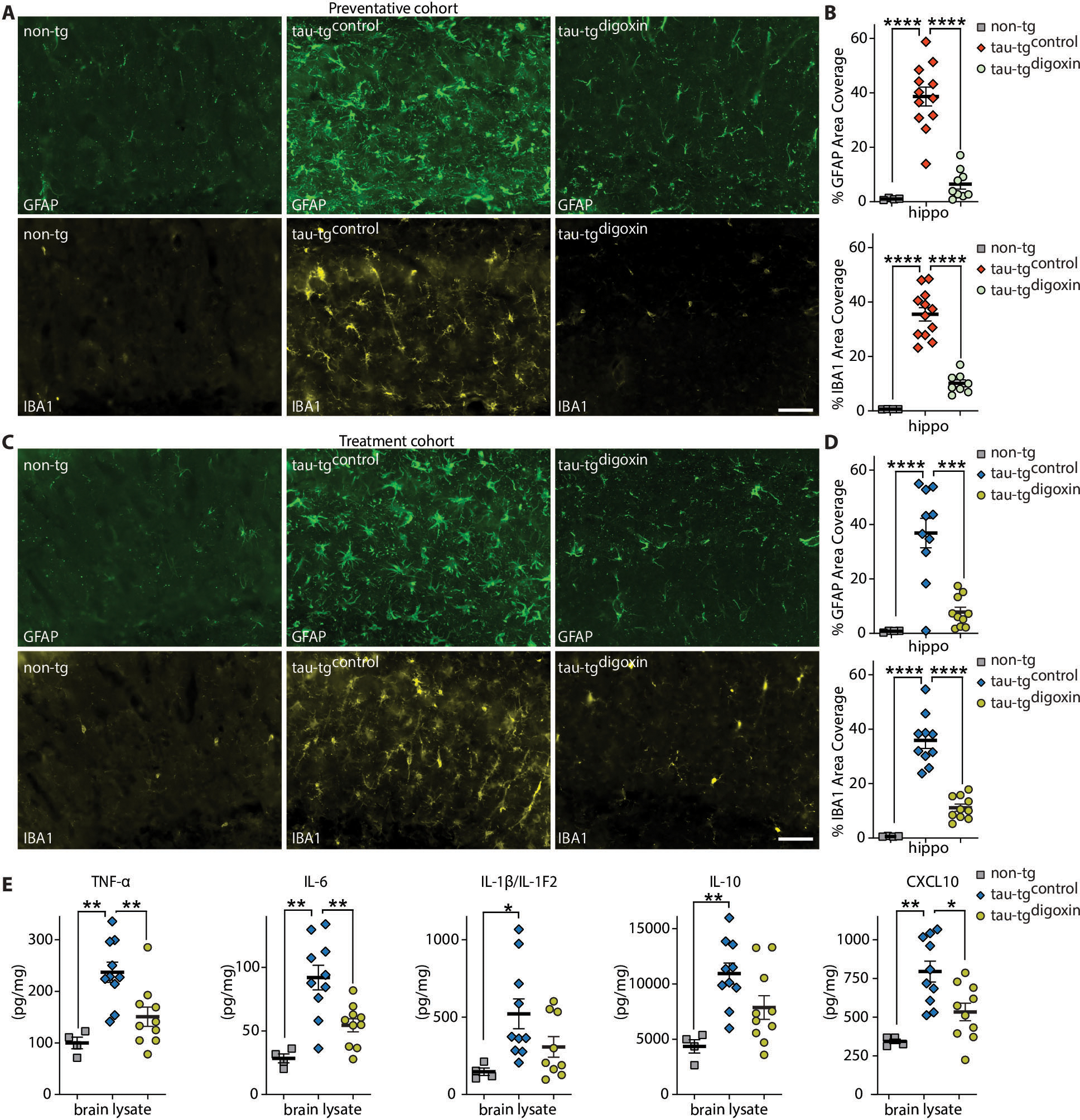
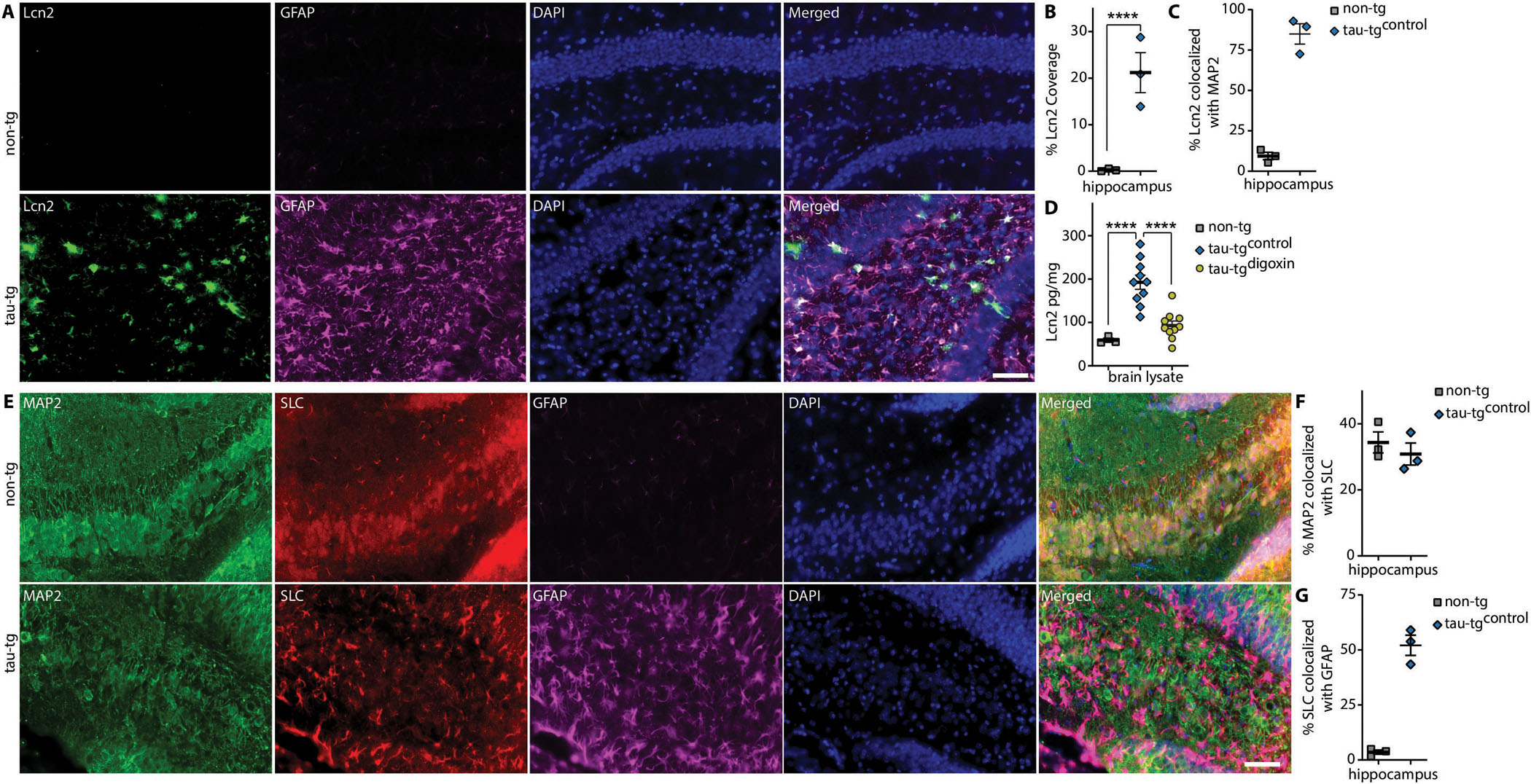
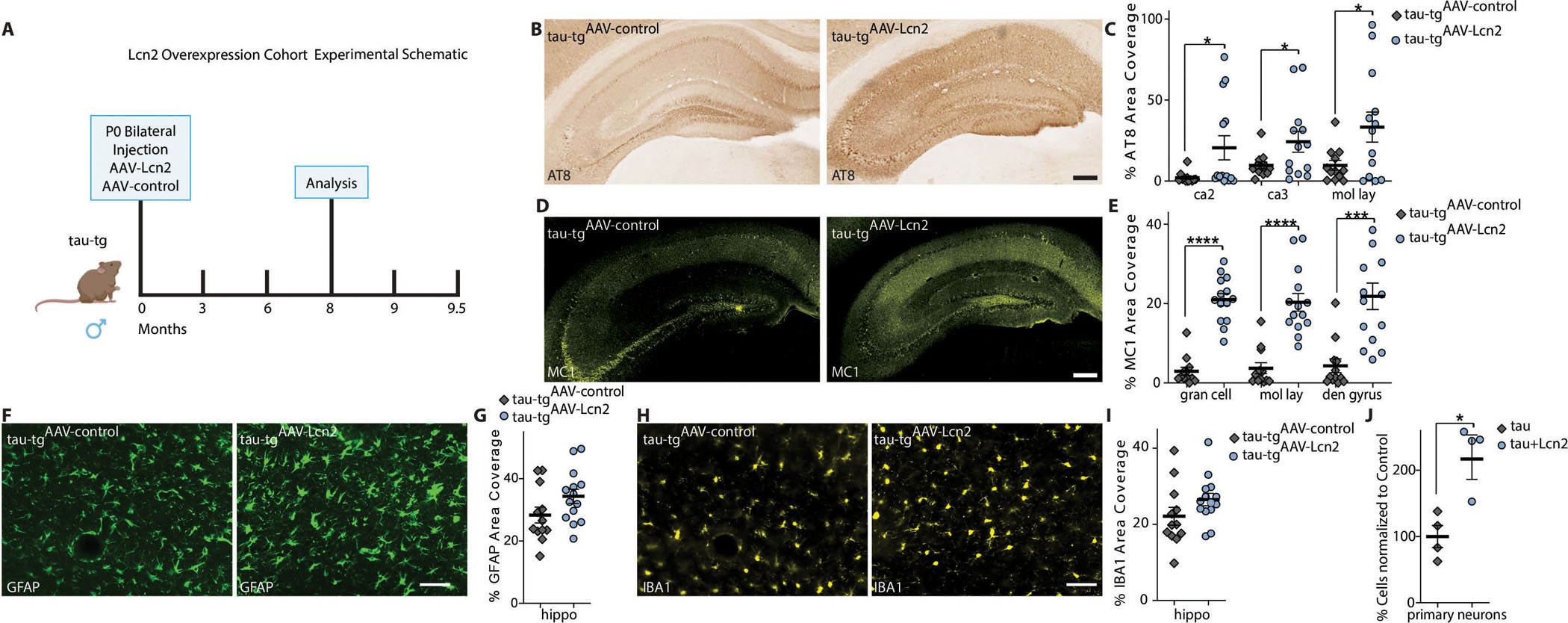
Alzheimer's disease (AD) is the most dominant form of dementia characterized by the deposition of extracellular amyloid plaques and intracellular neurofibrillary tau tangles (NFTs). In addition to these pathologies, an emerging pathophysiological mechanism that influences AD is neuroinflammation. Astrocytes are a vital type of glial cell that contribute to neuroinflammation, and reactive astrocytes, or astrogliosis, are a well-known pathological feature of AD. However, the mechanisms by which astrocytes contribute to the neurodegenerative process in AD have not been fully elucidated. Here, we showed that astrocytic α2-Na+/K+ adenosine triphosphatase (α2-NKA) is elevated in postmortem human brain tissue from AD and progressive nuclear palsy, a primary tauopathy. The increased astrocytic α2-NKA was also recapitulated in a mouse model of tauopathy. Pharmacological inhibition of α2-NKA robustly suppressed neuroinflammation and reduced brain atrophy. In addition, α2-NKA knockdown in tauopathy mice halted the accumulation of tau pathology. We also demonstrated that α2-NKA promoted tauopathy, in part, by regulating the proinflammatory protein lipocalin-2 (Lcn2). Overexpression of Lcn2 in tauopathy mice increased tau pathology, and prolonged Lcn2 exposure to primary neurons promoted tau uptake in vitro. These studies collectively highlight the contribution of reactive astrocytes to tau pathogenesis in mice and define α2-NKA as a major regulator of astrocytic-dependent neuroinflammation.
Conflict of interest statement
Figures
References
-
- Kidd M, Paired helical filaments in electron microscopy of Alzheimer’s disease. Nature 197, 192–193 (1963); published online EpubJan 12 ( - PubMed
-
- Joachim CL, Morris JH, Selkoe DJ, Kosik KS, Tau epitopes are incorporated into a range of lesions in Alzheimer’s disease. J Neuropathol Exp Neurol 46, 611–622 (1987); published online EpubNov ( - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 NS090934/NS/NINDS NIH HHS/United States
- RF1 NS090934/NS/NINDS NIH HHS/United States
- P50 AG016574/AG/NIA NIH HHS/United States
- P01 AG026276/AG/NIA NIH HHS/United States
- R01 AG032990/AG/NIA NIH HHS/United States
- R01 NS080820/NS/NINDS NIH HHS/United States
- P30 CA091842/CA/NCI NIH HHS/United States
- U01 AG046139/AG/NIA NIH HHS/United States
- R56 NS109007/NS/NINDS NIH HHS/United States
- P01 AG017216/AG/NIA NIH HHS/United States
- U01 AG006786/AG/NIA NIH HHS/United States
- U01 AI066576/AI/NIAID NIH HHS/United States
- P30 AG066444/AG/NIA NIH HHS/United States
- S10 RR027552/RR/NCRR NIH HHS/United States
- P01 AG003991/AG/NIA NIH HHS/United States
- P01 AG003949/AG/NIA NIH HHS/United States
- U24 NS072026/NS/NINDS NIH HHS/United States
- P30 AG019610/AG/NIA NIH HHS/United States
- P50 AG025711/AG/NIA NIH HHS/United States
- UL1 TR002345/TR/NCATS NIH HHS/United States
- R01 AG018023/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
