

2.4-Å structure of the double-ring Gemmatimonas phototrophica photosystem
- PMID: 35171663
- PMCID: PMC8849296
- DOI: 10.1126/sciadv.abk3139
2.4-Å structure of the double-ring Gemmatimonas phototrophica photosystem
Abstract
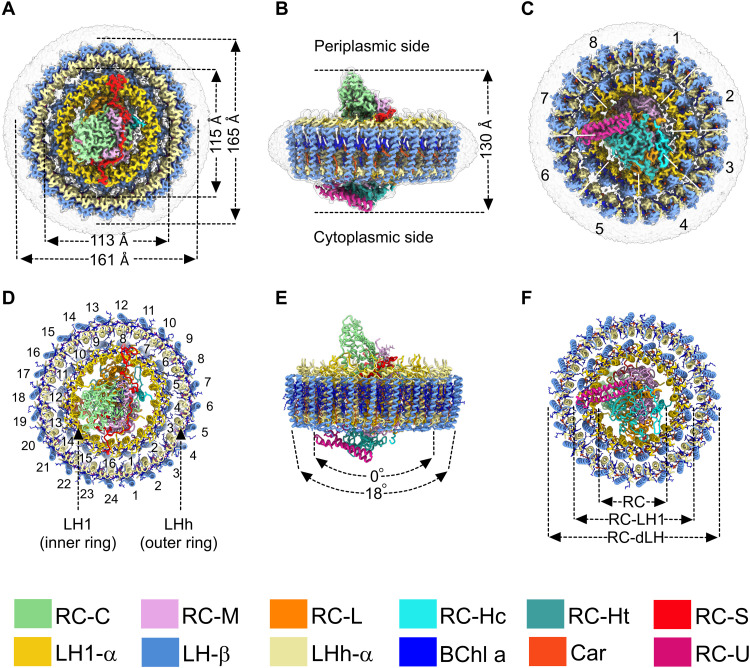
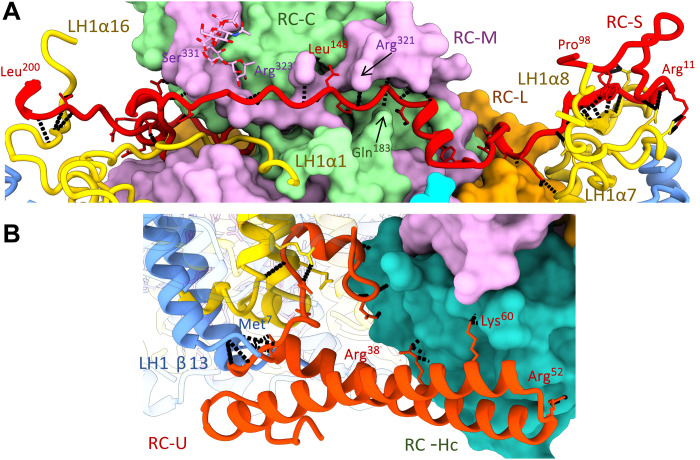
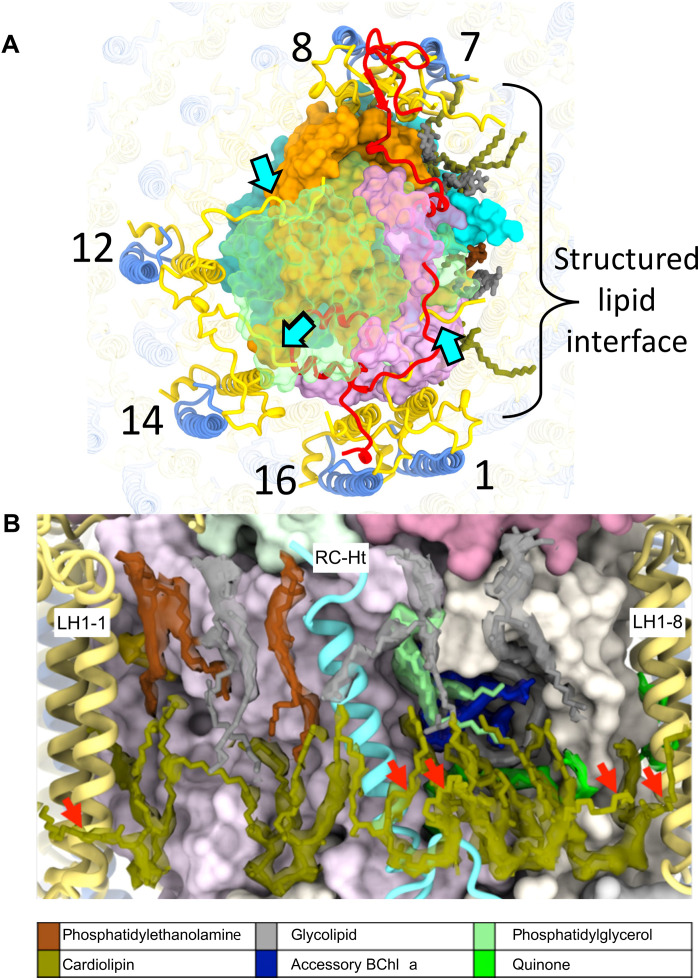
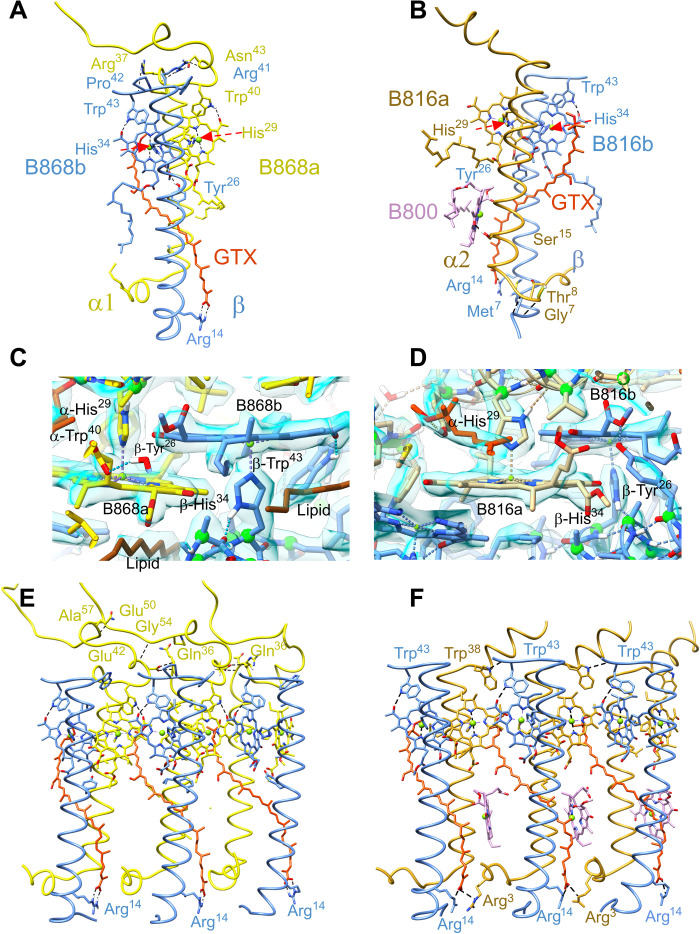
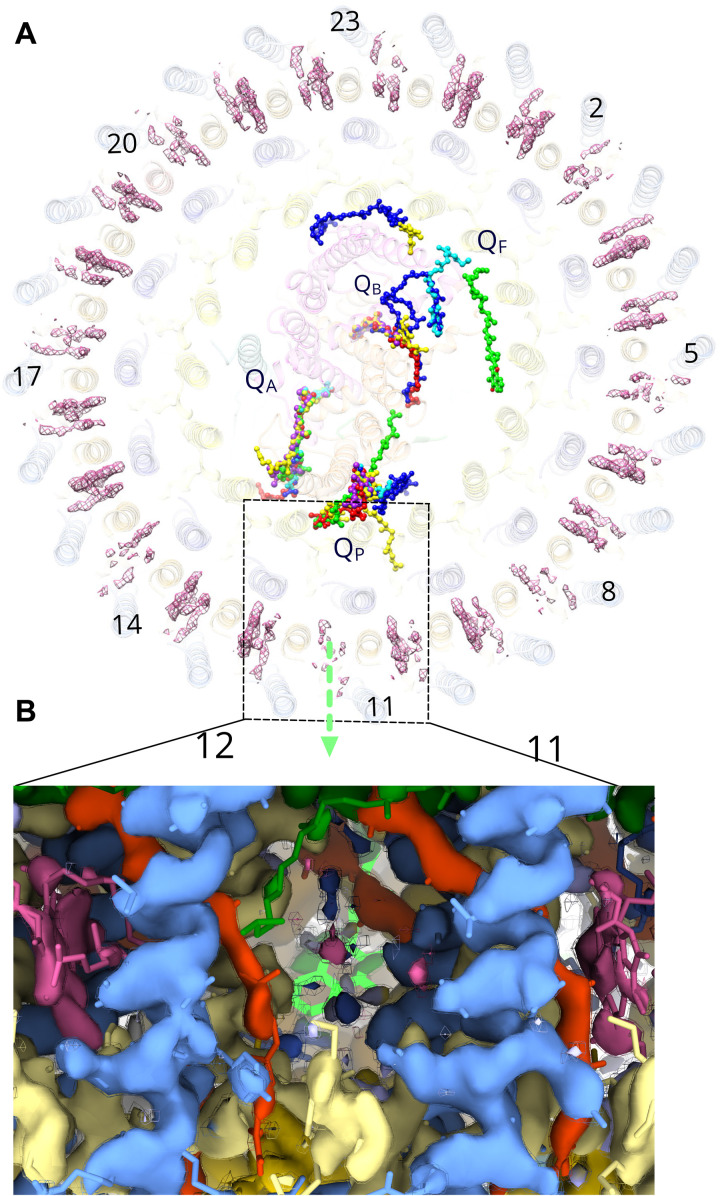
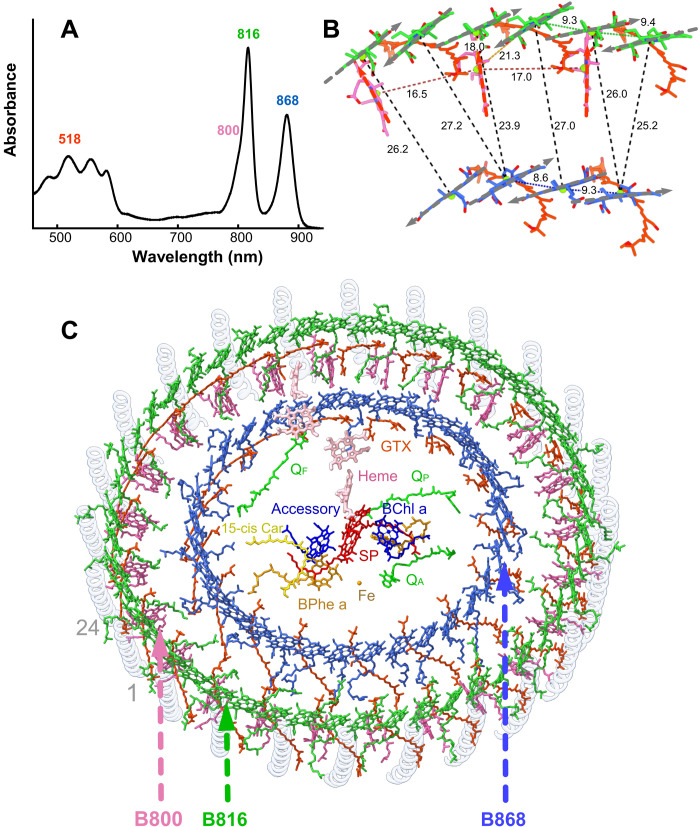
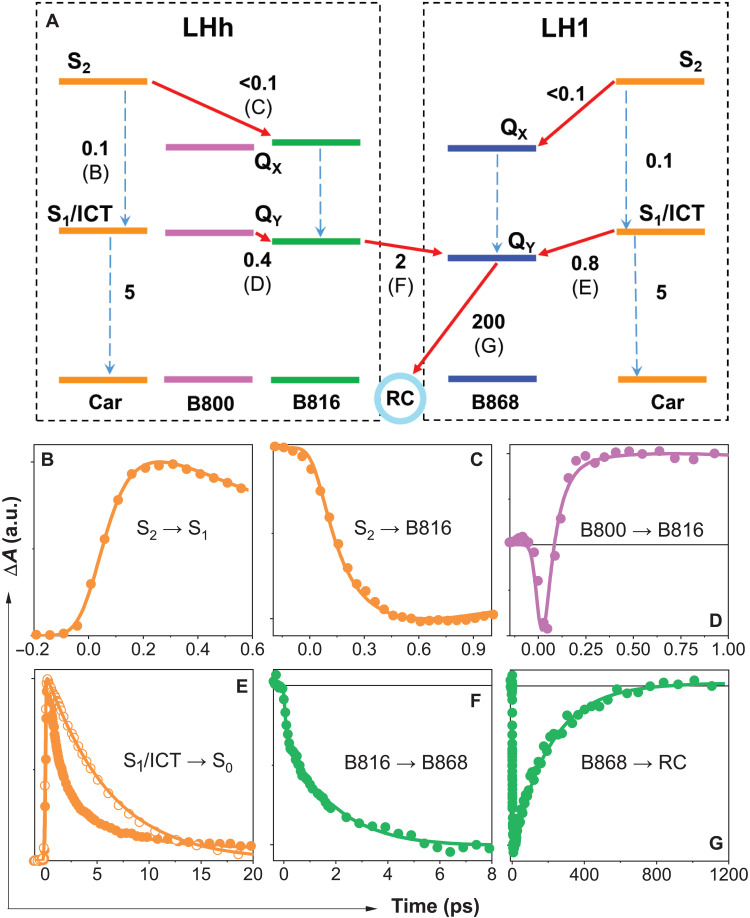
Phototrophic Gemmatimonadetes evolved the ability to use solar energy following horizontal transfer of photosynthesis-related genes from an ancient phototrophic proteobacterium. The electron cryo-microscopy structure of the Gemmatimonas phototrophica photosystem at 2.4 Å reveals a unique, double-ring complex. Two unique membrane-extrinsic polypeptides, RC-S and RC-U, hold the central type 2 reaction center (RC) within an inner 16-subunit light-harvesting 1 (LH1) ring, which is encircled by an outer 24-subunit antenna ring (LHh) that adds light-gathering capacity. Femtosecond kinetics reveal the flow of energy within the RC-dLH complex, from the outer LHh ring to LH1 and then to the RC. This structural and functional study shows that G. phototrophica has independently evolved its own compact, robust, and highly effective architecture for harvesting and trapping solar energy.
Figures
References
-
- R. E. Blankenship, Molecular Mechanisms of Photosynthesis (John Wiley & Sons, ed. 2, 2014), pp. 312.
-
- Hohmann-Marriott M. F., Blankenship R. E., Evolution of photosynthesis. Annu. Rev. Plant Biol. 62, 515–548 (2011). - PubMed
-
- Bryant D. A., Costas A. M., Maresca J. A., Chew A. G., Klatt C. G., Bateson M. M., Tallon L. J., Hostetler J., Nelson W. C., Heidelberg J. F., Ward D. M., Candidatus Chloracidobacterium thermophilum: An aerobic phototrophic Acidobacterium. Science 317, 523–526 (2007). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
