

Active anaerobic methane oxidation and sulfur disproportionation in the deep terrestrial subsurface
- PMID: 35173296
- PMCID: PMC9123182
- DOI: 10.1038/s41396-022-01207-w
Active anaerobic methane oxidation and sulfur disproportionation in the deep terrestrial subsurface
Abstract
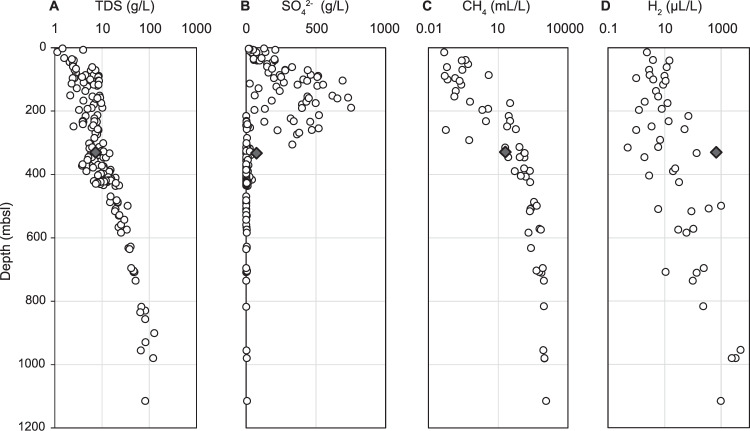
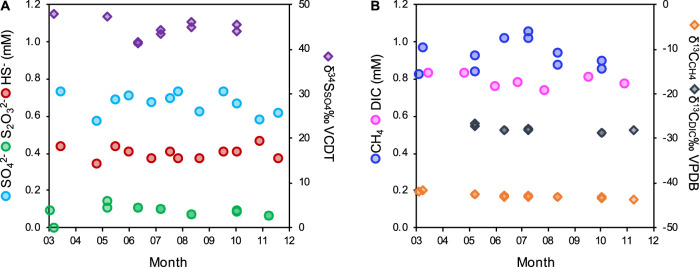
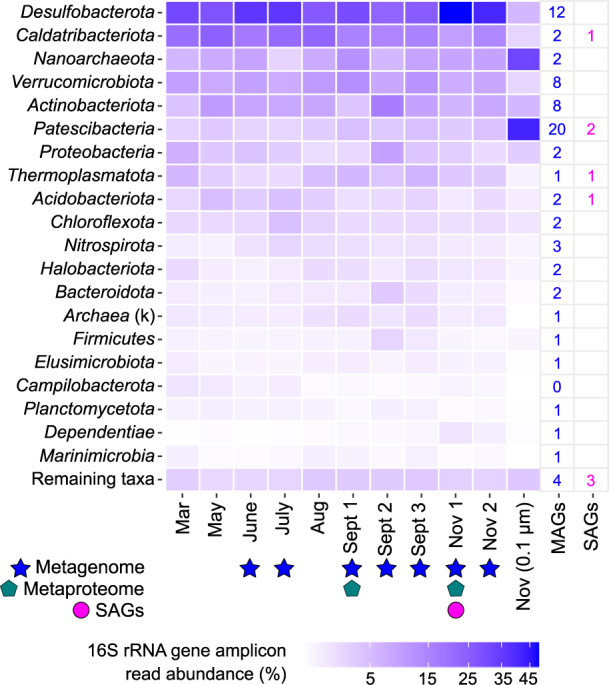
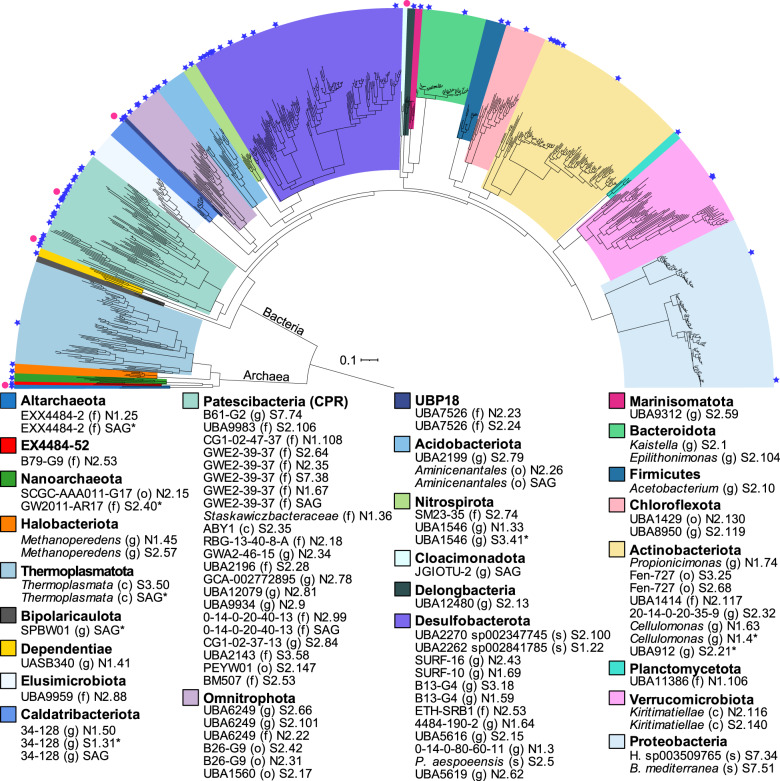
Microbial life is widespread in the terrestrial subsurface and present down to several kilometers depth, but the energy sources that fuel metabolism in deep oligotrophic and anoxic environments remain unclear. In the deep crystalline bedrock of the Fennoscandian Shield at Olkiluoto, Finland, opposing gradients of abiotic methane and ancient seawater-derived sulfate create a terrestrial sulfate-methane transition zone (SMTZ). We used chemical and isotopic data coupled to genome-resolved metaproteogenomics to demonstrate active life and, for the first time, provide direct evidence of active anaerobic oxidation of methane (AOM) in a deep terrestrial bedrock. Proteins from Methanoperedens (formerly ANME-2d) are readily identifiable despite the low abundance (≤1%) of this genus and confirm the occurrence of AOM. This finding is supported by 13C-depleted dissolved inorganic carbon. Proteins from Desulfocapsaceae and Desulfurivibrionaceae, in addition to 34S-enriched sulfate, suggest that these organisms use inorganic sulfur compounds as both electron donor and acceptor. Zerovalent sulfur in the groundwater may derive from abiotic rock interactions, or from a non-obligate syntrophy with Methanoperedens, potentially linking methane and sulfur cycles in Olkiluoto groundwater. Finally, putative episymbionts from the candidate phyla radiation (CPR) and DPANN archaea represented a significant diversity in the groundwater (26/84 genomes) with roles in sulfur and carbon cycling. Our results highlight AOM and sulfur disproportionation as active metabolisms and show that methane and sulfur fuel microbial activity in the deep terrestrial subsurface.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
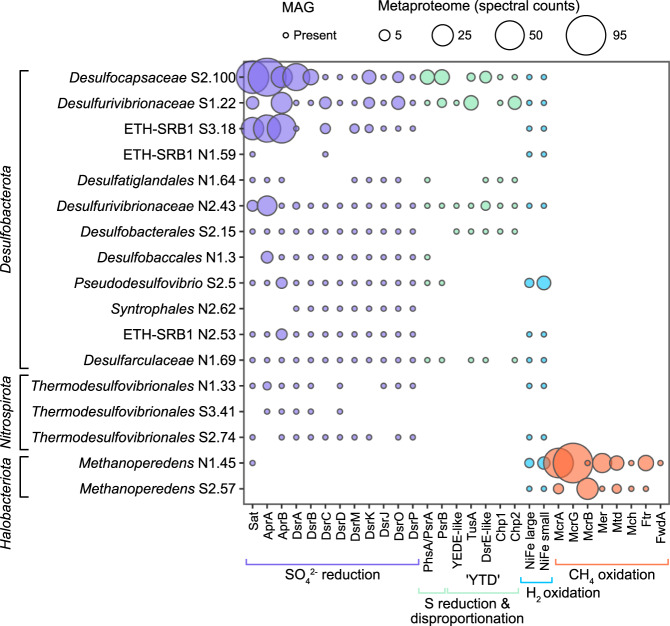
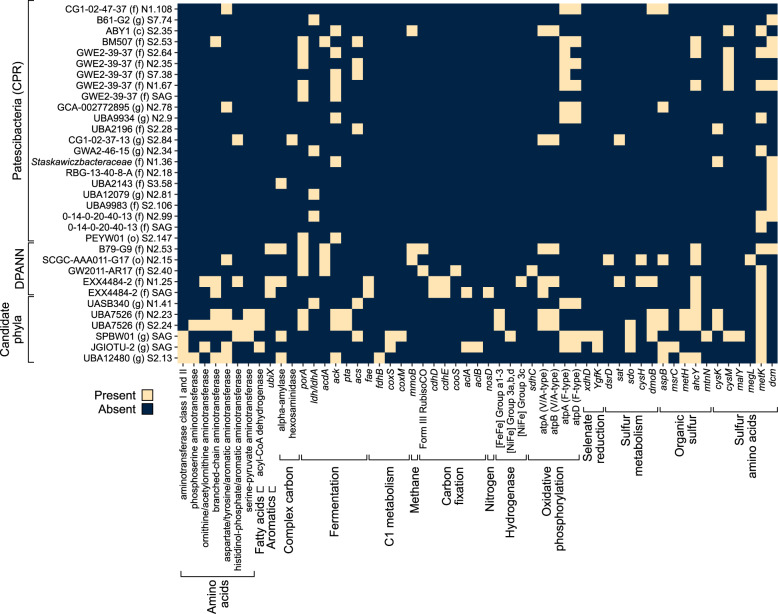
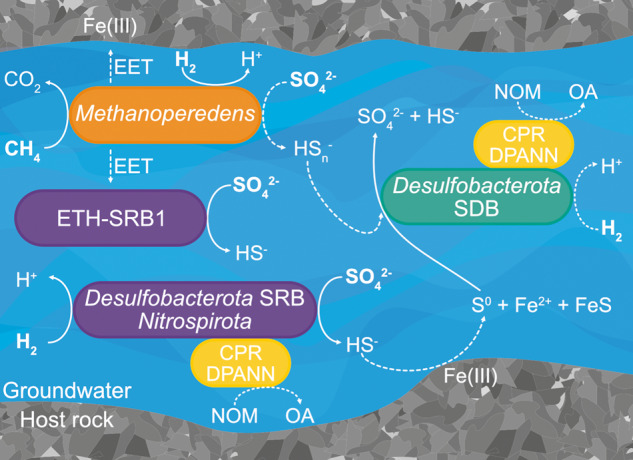
References
-
- Magnabosco C, Lin L-H, Dong H, Bomberg M, Ghiorse W, Stan-Lotter H, et al. The biomass and biodiversity of the continental subsurface. Nat Geosci. 2018;11:707–17.
-
- Hoehler TM, Jørgensen BB. Microbial life under extreme energy limitation. Nat Rev Microbiol. 2013;11:83–94. - PubMed
