

Assessing Mitochondrial DNA Release into the Cytosol and Subsequent Activation of Innate Immune-related Pathways in Mammalian Cells
- PMID: 35175686
- PMCID: PMC8986093
- DOI: 10.1002/cpz1.372
Assessing Mitochondrial DNA Release into the Cytosol and Subsequent Activation of Innate Immune-related Pathways in Mammalian Cells
Erratum in
-
Correction: Assessing Mitochondrial DNA Release into the Cytosol and Subsequent Activation of Innate Immune-related Pathways in Mammalian Cells.Curr Protoc. 2024 Jul;4(7):e1106. doi: 10.1002/cpz1.1106. Curr Protoc. 2024. PMID: 38967479 No abstract available.
Abstract
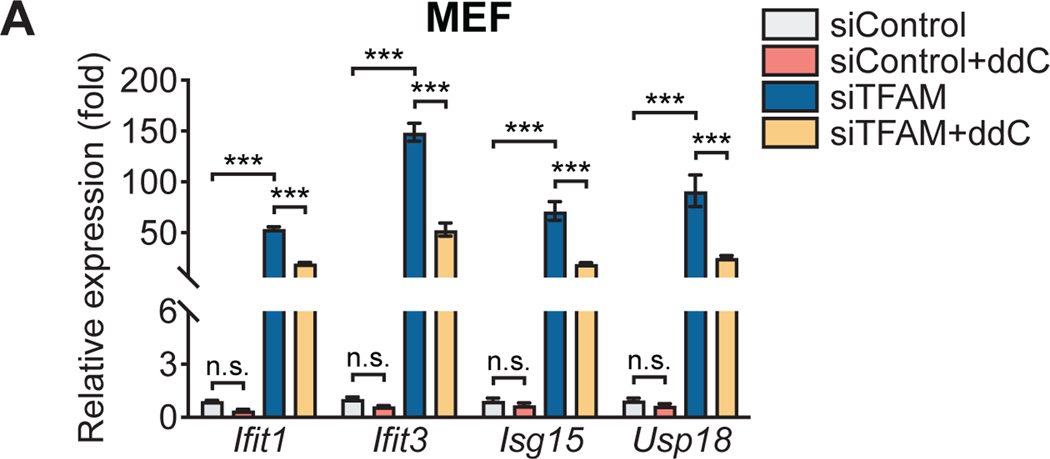
Mitochondria have emerged as key drivers of mammalian innate immune responses, functioning as signaling hubs to trigger inflammation and orchestrating metabolic switches required for phagocyte activation. Mitochondria also contain damage-associated molecular patterns (DAMPs), molecules that share similarity with pathogen-associated molecular patterns (PAMPs) and can engage innate immune sensors to drive inflammation. The aberrant release of mitochondrial DAMPs during cellular stress and injury is an increasingly recognized trigger of inflammatory responses in human diseases. Mitochondrial DNA (mtDNA) is a particularly potent DAMP that engages multiple innate immune sensors, although mounting evidence suggests that cytosolic mtDNA is primarily detected via the cyclic GMP-AMP synthase-stimulator of interferon genes (cGAS-STING) pathway. cGAS and STING are widely expressed in mammalian cells and serve as key regulators of type I interferon and cytokine expression in both infectious and inflammatory diseases. Despite growing roles for the mtDNA-cGAS-STING axis in human disease, assays to quantify mtDNA release into the cytosol and approaches to link mtDNA to cGAS-STING signaling are not standardized, which increases the possibility for experimental artifacts and misinterpretation of data. Here, we present a series of protocols for assaying the release of mtDNA into the cytosol and subsequent activation of innate immune signaling in mammalian cells. We highlight genetic and pharmacological approaches to induce and inhibit mtDNA release from mitochondria. We also describe immunofluorescence microscopy and cellular fractionation assays to visualize morphological changes in mtDNA and quantify mtDNA accumulation in the cytosol. Finally, we include protocols to examine mtDNA-dependent cGAS-STING activation by RT-qPCR and western blotting. These methods can be performed with standard laboratory equipment and are highly adaptable to a wide range of mammalian cell types. They will permit researchers working across the spectrum of biological and biomedical sciences to accurately and reproducibly measure cytosolic mtDNA release and resulting innate immune responses. © 2022 Wiley Periodicals LLC. Basic Protocol 1: siRNA-mediated knockdown of TFAM to induce mtDNA instability, cytosolic release, and activation of the cGAS-STING pathway Alternate Protocol: Pharmacological induction of mtDNA release and cGAS-STING activation using ABT-737 and Q-VD-OPH Basic Protocol 2: Isolation and quantitation of DNA from cytosolic, mitochondrial, and nuclear fractions Basic Protocol 3: Pharmacological inhibition of mtDNA replication and release.
Keywords: STING; cGAS; innate immunity; mitochondria; mitochondrial DNA.
© 2022 Wiley Periodicals LLC.
Conflict of interest statement
CONFLICT OF INTEREST STATEMENT:
The authors declare that they have no competing interests.
Figures





References
-
- Akira S, and Takeda K 2004. Toll-like receptor signalling. Nature Reviews Immunology 4:499–511. - PubMed
-
- Andersen CL, Jensen JL, and Ørntoft TF 2004. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Research 64:5245–5250. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
