

RelB upregulates PD-L1 and exacerbates prostate cancer immune evasion
- PMID: 35177112
- PMCID: PMC8851785
- DOI: 10.1186/s13046-022-02243-2
RelB upregulates PD-L1 and exacerbates prostate cancer immune evasion
Abstract
Background: The interaction between programmed death receptor (PD-1) and its ligand (PD-L1) is essential for suppressing activated T-lymphocytes. However, the precise mechanisms underlying PD-L1 overexpression in tumours have yet to be fully elucidated. Here, we describe that RelB participates in the immune evasion of prostate cancer (PCa) via cis/trans transcriptional upregulation of PD-L1.
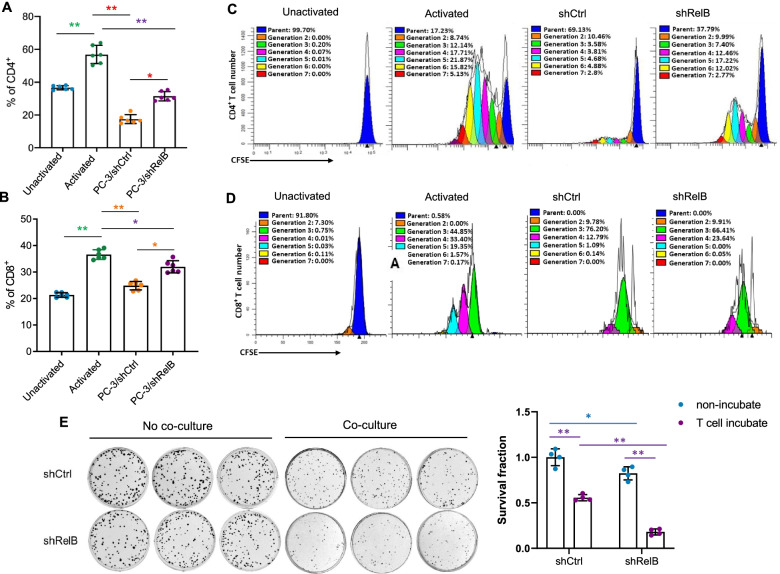
Methods: Based on transcriptome results, RelB was manipulated in multiple human and murine PCa cell lines. Activated CD4+ and CD8+ T cells were cocultured with PCa cells with different levels of RelB to examine the effect of tumourous RelB on T cell immunity. Male mice were injected with murine PCa cells to validate the effect of RelB on the PD-1/PD-L1-mediated immune checkpoint using both tumour growth and metastatic experimental models.
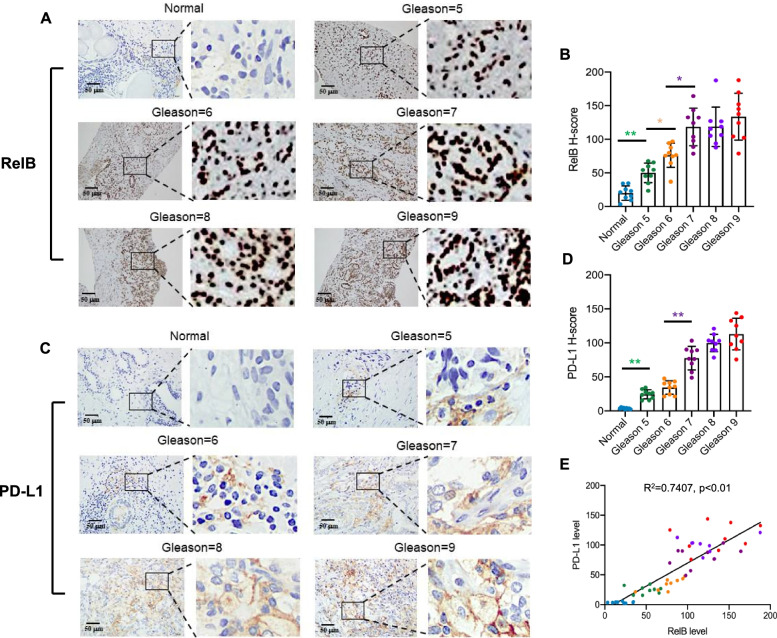
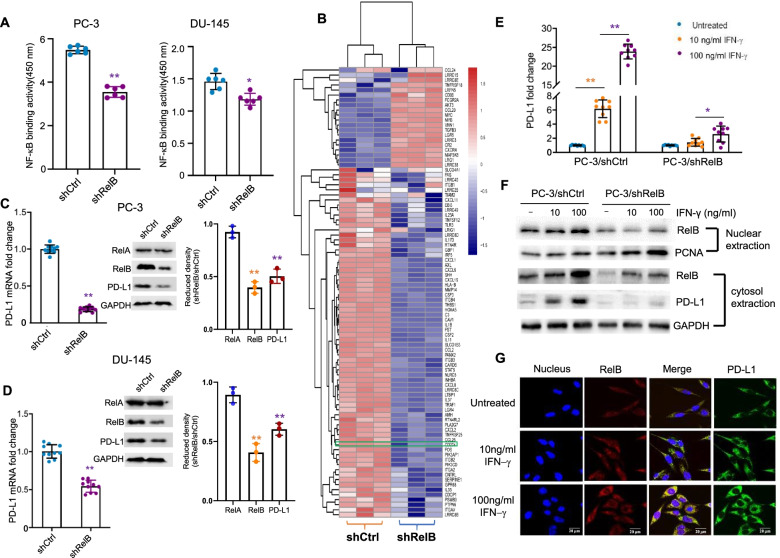
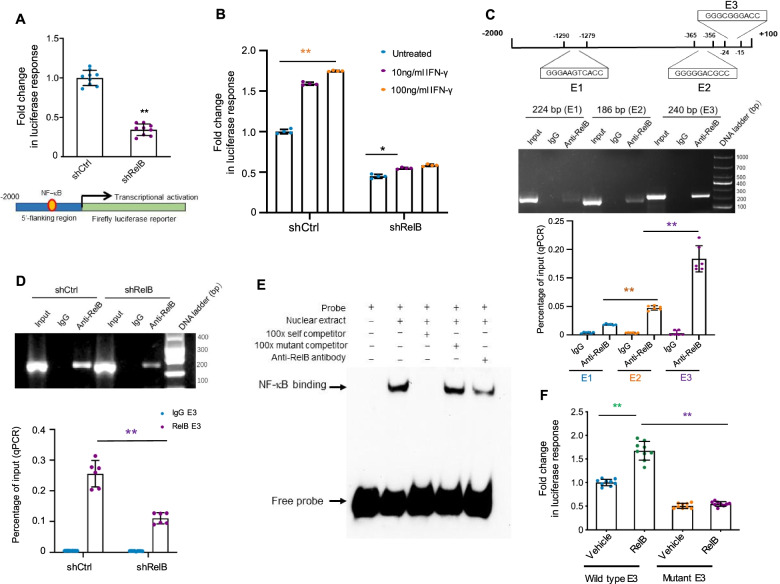
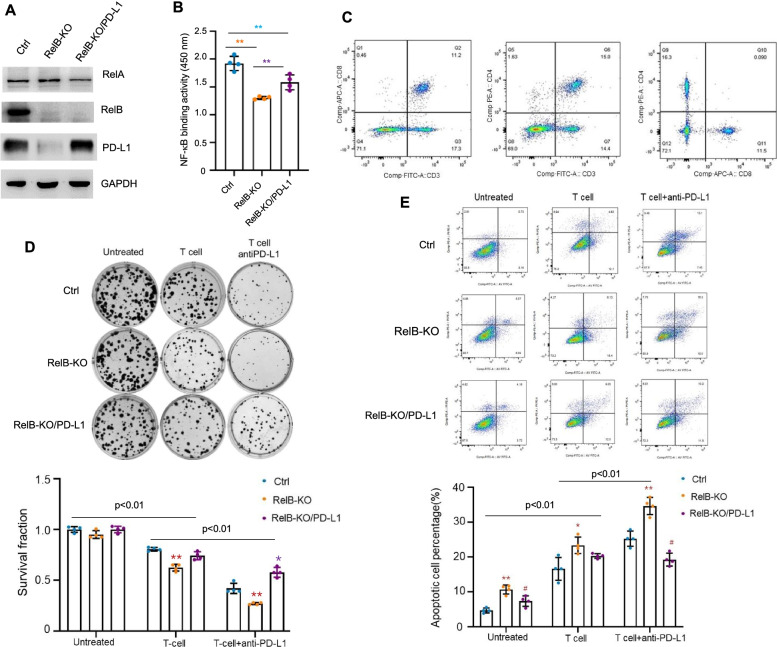
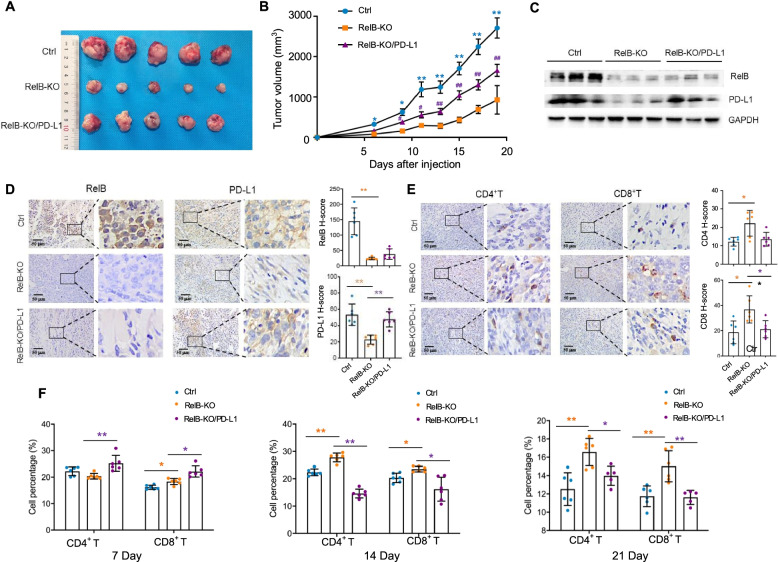
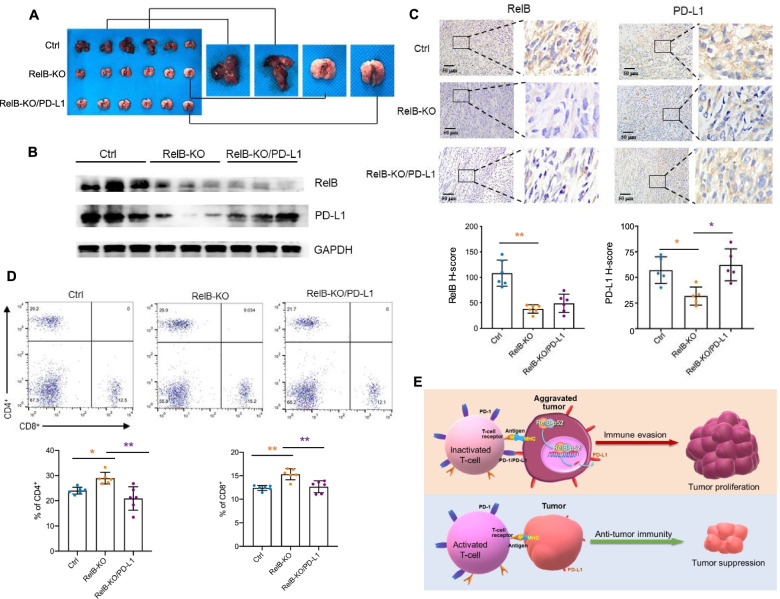
Results: PD-L1 is uniquely expressed at a high level in PCa with high constitutive RelB and correlates with the patients' Gleason scores. Indeed, a high level of PD-L1 is associated with RelB nuclear translocation in AR-negative aggressive PCa cells. Conversely, the silencing of RelB in advanced PCa cells resulted in reduced PD-L1 expression and enhanced susceptibility of PCa cells to the T cell immune response in vitro and in vivo. Mechanistically, a proximal NF-κB enhancer element was identified in the core promoter region of the human CD274 gene, which is responsible for RelB-mediated PD-L1 transcriptional activation. This finding provides an informative insight into immune checkpoint blockade by administering RelB within the tumour microenvironment.
Conclusion: This study deciphers the molecular mechanism by which tumourous RelB contributes to immune evasion by inhibiting T cell immunity via the amplification of the PD-L1/PD-1-mediated immune checkpoint.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Insight into prostate cancer osteolytic metastasis by RelB coordination of IL-8 and S100A4.Clin Transl Med. 2024 Oct;14(10):e70058. doi: 10.1002/ctm2.70058. Clin Transl Med. 2024. PMID: 39415352 Free PMC article.
-
Inverse relationship between PSA and IL-8 in prostate cancer: an insight into a NF-κB-mediated mechanism.PLoS One. 2012;7(3):e32905. doi: 10.1371/journal.pone.0032905. Epub 2012 Mar 5. PLoS One. 2012. PMID: 22403723 Free PMC article.
-
NDR1 mediates PD-L1 deubiquitination to promote prostate cancer immune escape via USP10.Cell Commun Signal. 2024 Sep 3;22(1):429. doi: 10.1186/s12964-024-01805-5. Cell Commun Signal. 2024. PMID: 39227807 Free PMC article.
-
Vitamin D receptor-mediated suppression of RelB in antigen presenting cells: a paradigm for ligand-augmented negative transcriptional regulation.Arch Biochem Biophys. 2007 Apr 15;460(2):218-26. doi: 10.1016/j.abb.2007.01.034. Epub 2007 Feb 23. Arch Biochem Biophys. 2007. PMID: 17367745 Free PMC article. Review.
-
Molecular panorama of therapy resistance in prostate cancer: a pre-clinical and bioinformatics analysis for clinical translation.Cancer Metastasis Rev. 2024 Mar;43(1):229-260. doi: 10.1007/s10555-024-10168-9. Epub 2024 Feb 19. Cancer Metastasis Rev. 2024. PMID: 38374496 Review.
Cited by
-
Insight into prostate cancer osteolytic metastasis by RelB coordination of IL-8 and S100A4.Clin Transl Med. 2024 Oct;14(10):e70058. doi: 10.1002/ctm2.70058. Clin Transl Med. 2024. PMID: 39415352 Free PMC article.
-
Current insight into the regulation of PD-L1 in cancer.Exp Hematol Oncol. 2022 Jul 30;11(1):44. doi: 10.1186/s40164-022-00297-8. Exp Hematol Oncol. 2022. PMID: 35907881 Free PMC article. Review.
-
Hexavalent chromium exposure activates the non-canonical nuclear factor kappa B pathway to promote immune checkpoint protein programmed death-ligand 1 expression and lung carcinogenesis.Cancer Lett. 2024 May 1;589:216827. doi: 10.1016/j.canlet.2024.216827. Epub 2024 Mar 23. Cancer Lett. 2024. PMID: 38527692 Free PMC article.
-
Inhibitory immune checkpoints suppress the surveillance of senescent cells promoting their accumulation with aging and in age-related diseases.Biogerontology. 2024 Oct;25(5):749-773. doi: 10.1007/s10522-024-10114-w. Epub 2024 Jul 1. Biogerontology. 2024. PMID: 38954358 Free PMC article. Review.
-
NF-κB in biology and targeted therapy: new insights and translational implications.Signal Transduct Target Ther. 2024 Mar 4;9(1):53. doi: 10.1038/s41392-024-01757-9. Signal Transduct Target Ther. 2024. PMID: 38433280 Free PMC article. Review.
References
-
- Culp MB, Soerjomataram I, Efstathiou JA, Bray F, Jemal A. Recent global patterns in prostate Cancer incidence and mortality rates. Eur Urol. 2020;77:38–52. - PubMed
-
- Miller KD, Nogueira L, Mariotto AB, Rowland JH, Yabroff KR, Alfano CM, Jemal A, Kramer JL, Siegel RL. Cancer treatment and survivorship statistics, 2019. CA Cancer J Clin. 2019;69:363–385. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
