

Indole-3-Acetic Acid Alters Intestinal Microbiota and Alleviates Ankylosing Spondylitis in Mice
- PMID: 35185872
- PMCID: PMC8854167
- DOI: 10.3389/fimmu.2022.762580
Indole-3-Acetic Acid Alters Intestinal Microbiota and Alleviates Ankylosing Spondylitis in Mice
Abstract
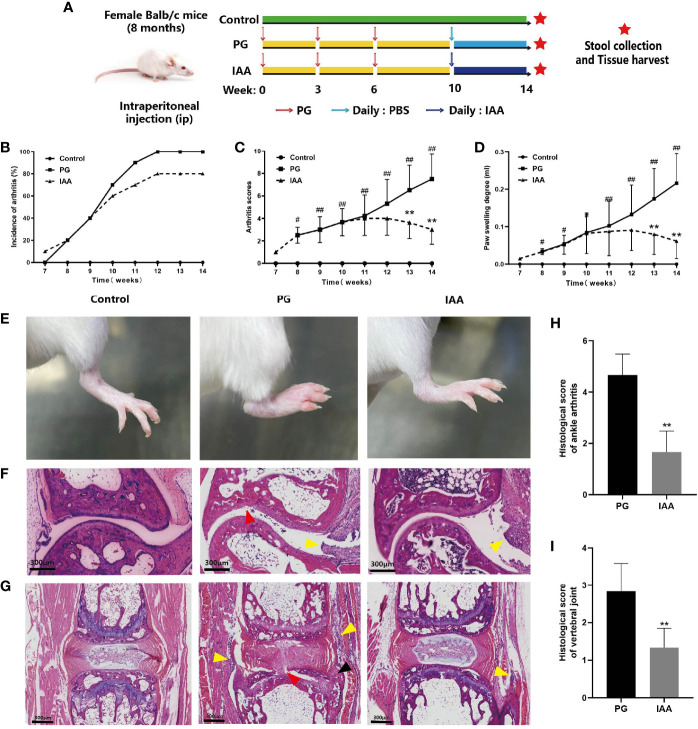
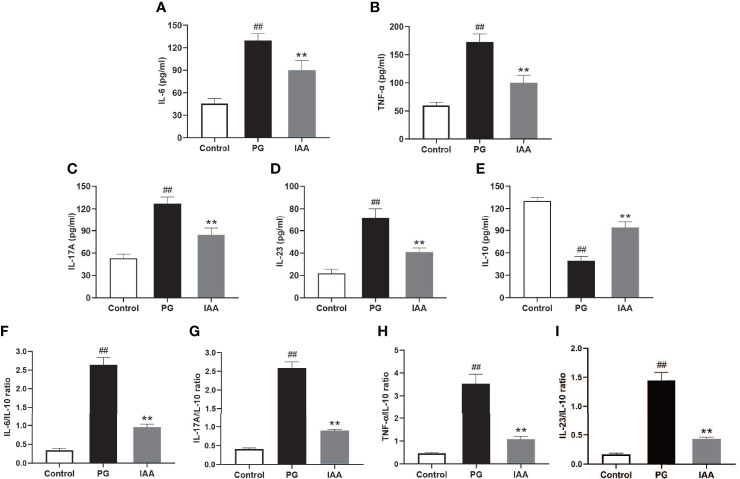
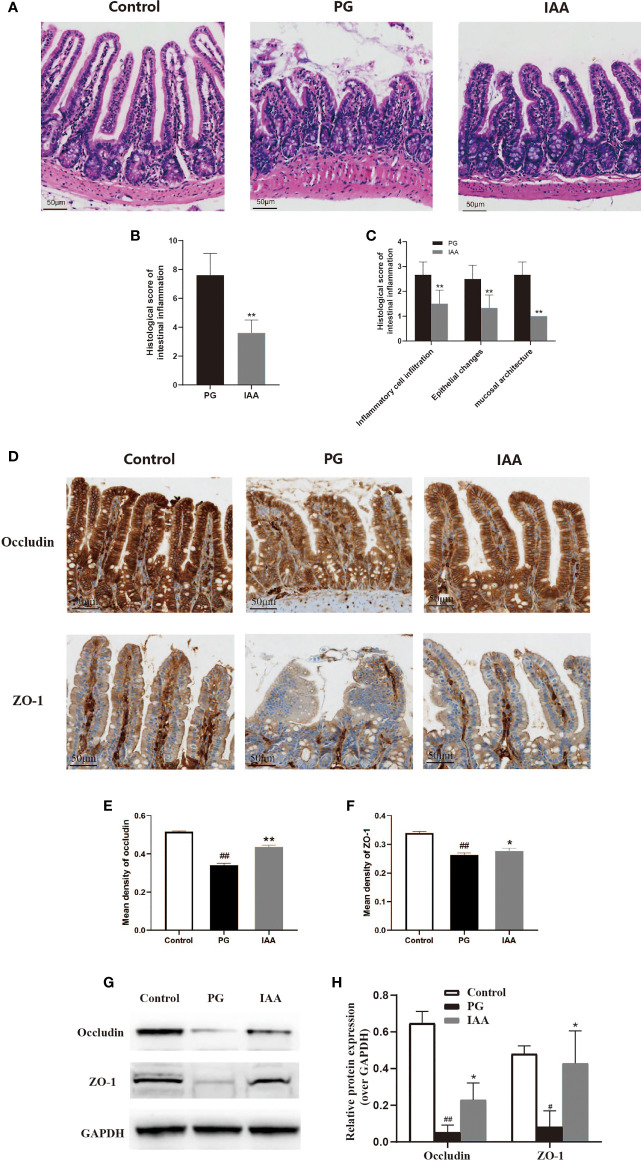
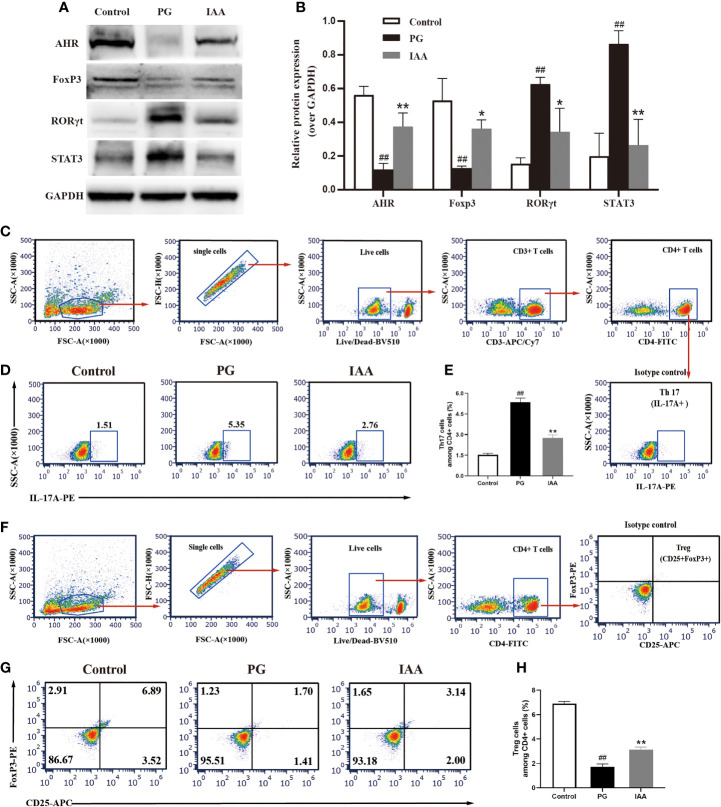
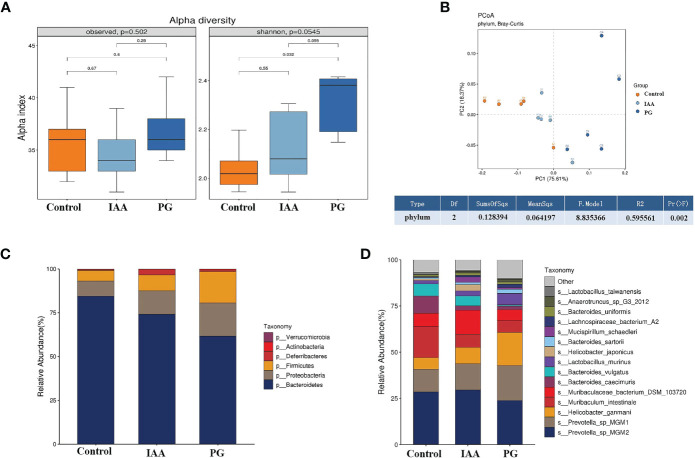
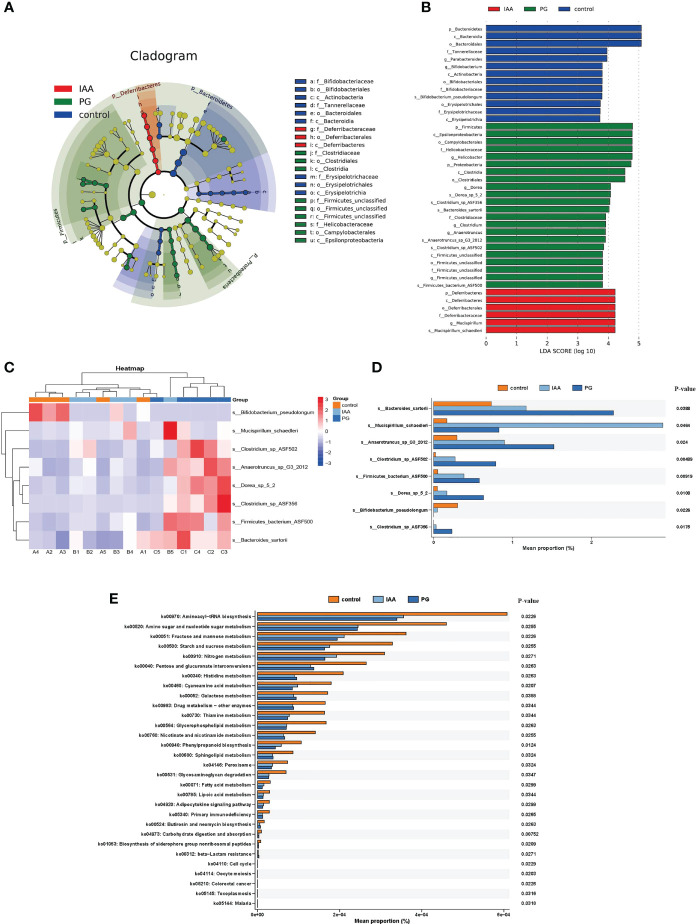
Ankylosing spondylitis (AS) is a systemic, chronic, and inflammatory autoimmune disease associated with the disorder of intestinal microbiota. Unfortunately, effective therapies for AS are lacking. Recent evidence has indicated that indole-3-acetic acid (IAA), an important microbial tryptophan metabolite, can modulate intestinal homeostasis and suppress inflammatory responses. However, reports have not examined the in vivo protective effects of IAA against AS. In this study, we investigated the protective effects and underlying mechanisms through which IAA acts against AS. We constructed a proteoglycan (PG)-induced AS mouse model and administered IAA (50 mg/kg body weight) by intraperitoneal injection daily for 4 weeks. The effects of IAA on AS mice were evaluated by examining disease severity, intestinal barrier function, aryl hydrocarbon receptor (AhR) pathway, T-helper 17 (Th17)/T regulatory (Treg) balance, and inflammatory cytokine levels. The intestinal microbiota compositions were profiled through whole-genome sequencing. We observed that IAA decreased the incidence and severity of AS in mice, inhibited the production of pro-inflammatory cytokines (tumor necrosis factor α [TNF-α], interleukin [IL]-6, IL-17A, and IL-23), promoted the production of the anti-inflammatory cytokine IL-10, and reduced the ratios of pro-/anti- inflammatory cytokines. IAA ameliorated pathological changes in the ileum and improved intestinal mucosal barrier function. IAA also activated the AhR pathway, upregulated the transcription factor forehead box protein P3 (FoxP3) and increased Treg cells, and downregulated the transcription factors retinoic acid receptor-related orphan receptor gamma t (RORγt) and signal transducer and activator of transcription 3 (STAT3) and decreased Th17 cells. Furthermore, IAA altered the composition of the intestinal microbiota composition by increasing Bacteroides and decreasing Proteobacteria and Firmicutes, in addition to increasing the abundances of Bifidobacterium pseudolongum and Mucispirillum schaedleri. In conclusion, IAA exerted several protective effects against PG-induced AS in mice, which was mediated by the restoration of balance among the intestinal microbial community, activating the AhR pathway, and inhibiting inflammation. IAA might represent a novel therapeutic approach for AS.
Keywords: ankylosing spondylitis; aryl hydrocarbon receptor; indole-3-acetic acid; inflammatory response; intestinal microbiota.
Copyright © 2022 Shen, Yang, You, Chen, Su, Cui, Wang, Zhang, Liu, Zhou and Lu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous