

Enhancer retargeting of CDX2 and UBTF::ATXN7L3 define a subtype of high-risk B-progenitor acute lymphoblastic leukemia
- PMID: 35192684
- PMCID: PMC9203703
- DOI: 10.1182/blood.2022015444
Enhancer retargeting of CDX2 and UBTF::ATXN7L3 define a subtype of high-risk B-progenitor acute lymphoblastic leukemia
Abstract
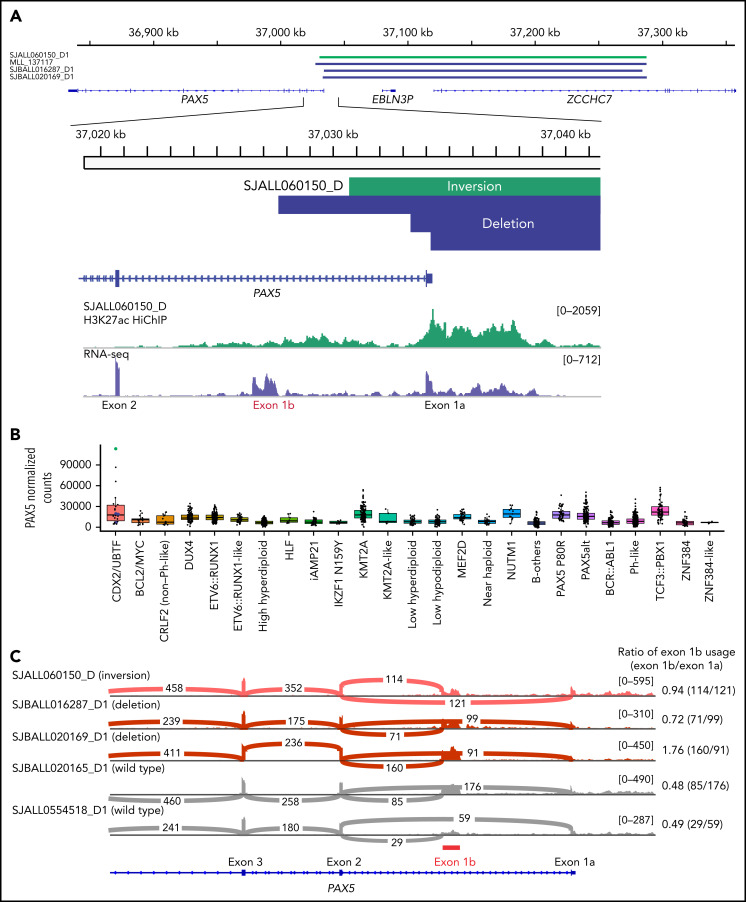
Transcriptome sequencing has identified multiple subtypes of B-progenitor acute lymphoblastic leukemia (B-ALL) of prognostic significance, but a minority of cases lack a known genetic driver. Here, we used integrated whole-genome (WGS) and -transcriptome sequencing (RNA-seq), enhancer mapping, and chromatin topology analysis to identify previously unrecognized genomic drivers in B-ALL. Newly diagnosed (n = 3221) and relapsed (n = 177) B-ALL cases with tumor RNA-seq were studied. WGS was performed to detect mutations, structural variants, and copy number alterations. Integrated analysis of histone 3 lysine 27 acetylation and chromatin looping was performed using HiChIP. We identified a subset of 17 newly diagnosed and 5 relapsed B-ALL cases with a distinct gene expression profile and 2 universal and unique genomic alterations resulting from aberrant recombination-activating gene activation: a focal deletion downstream of PAN3 at 13q12.2 resulting in CDX2 deregulation by the PAN3 enhancer and a focal deletion of exons 18-21 of UBTF at 17q21.31 resulting in a chimeric fusion, UBTF::ATXN7L3. A subset of cases also had rearrangement and increased expression of the PAX5 gene, which is otherwise uncommon in B-ALL. Patients were more commonly female and young adult with median age 35 (range,12-70 years). The immunophenotype was characterized by CD10 negativity and immunoglobulin M positivity. Among 16 patients with known clinical response, 9 (56.3%) had high-risk features including relapse (n = 4) or minimal residual disease >1% at the end of remission induction (n = 5). CDX2-deregulated, UBTF::ATXN7L3 rearranged (CDX2/UBTF) B-ALL is a high-risk subtype of leukemia in young adults for which novel therapeutic approaches are required.
© 2022 by The American Society of Hematology.
Figures
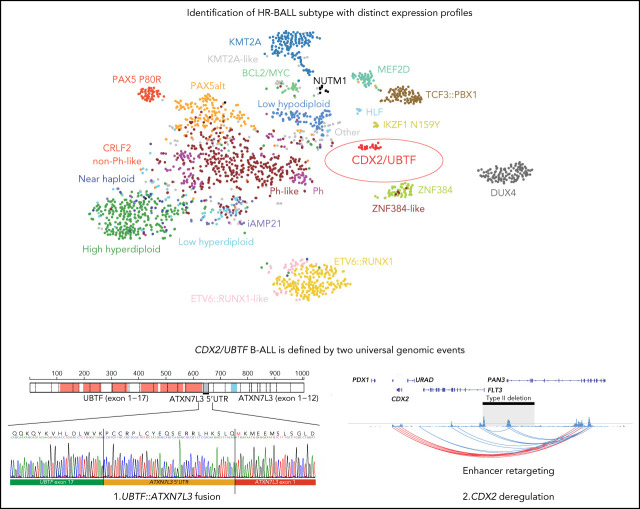
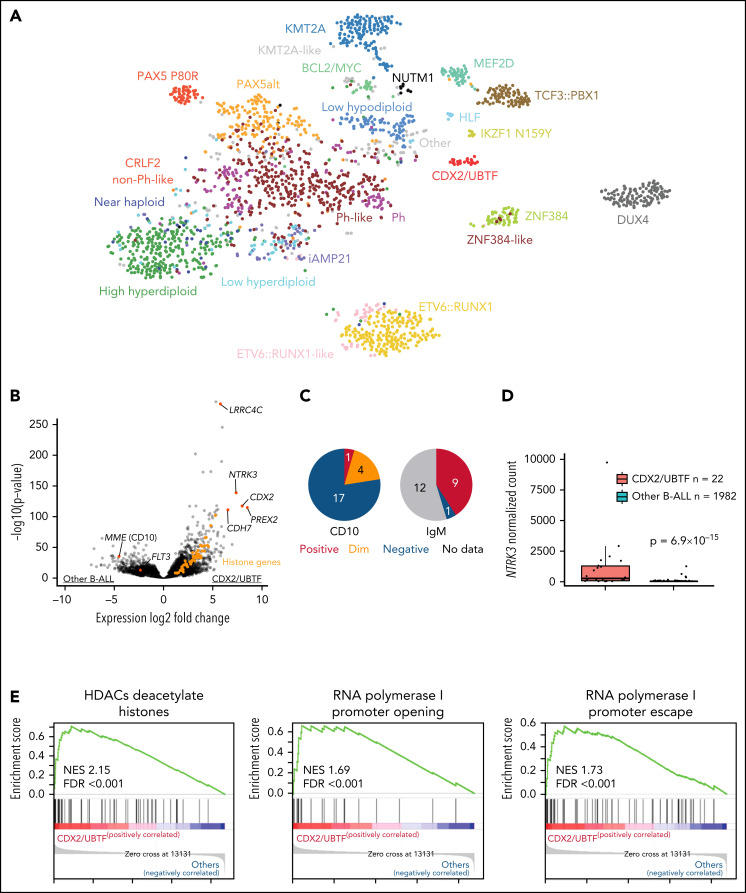
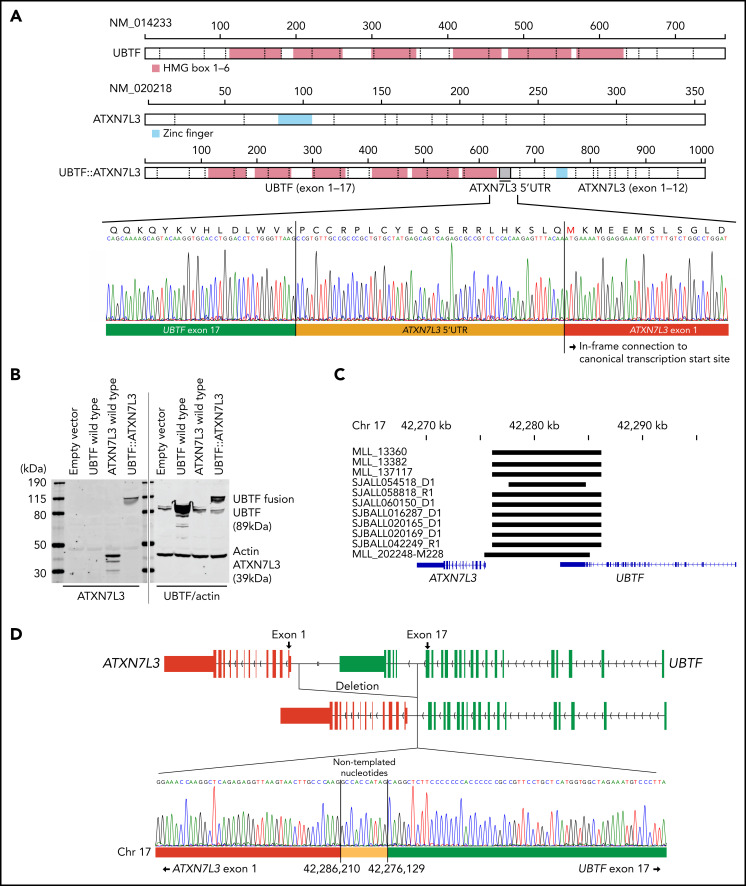
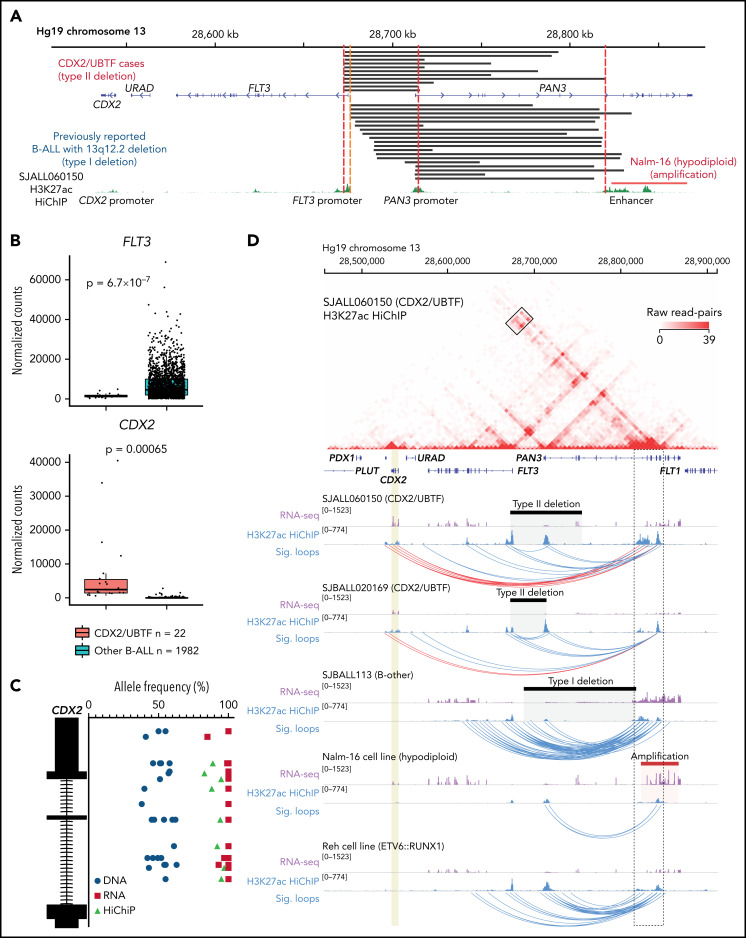
Comment in
-
Why B(-)other? About the gap of unknowns in ALL.Blood. 2022 Jun 16;139(24):3455-3457. doi: 10.1182/blood.2022015993. Blood. 2022. PMID: 35708724 Free PMC article. No abstract available.
References
-
- Hunger SP, Mullighan CG. Acute lymphoblastic leukemia in children. N Engl J Med. 2015;373(16):1541-1552. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
