

Systematic benchmark of state-of-the-art variant calling pipelines identifies major factors affecting accuracy of coding sequence variant discovery
- PMID: 35193511
- PMCID: PMC8862519
- DOI: 10.1186/s12864-022-08365-3
Systematic benchmark of state-of-the-art variant calling pipelines identifies major factors affecting accuracy of coding sequence variant discovery
Abstract
Background: Accurate variant detection in the coding regions of the human genome is a key requirement for molecular diagnostics of Mendelian disorders. Efficiency of variant discovery from next-generation sequencing (NGS) data depends on multiple factors, including reproducible coverage biases of NGS methods and the performance of read alignment and variant calling software. Although variant caller benchmarks are published constantly, no previous publications have leveraged the full extent of available gold standard whole-genome (WGS) and whole-exome (WES) sequencing datasets.
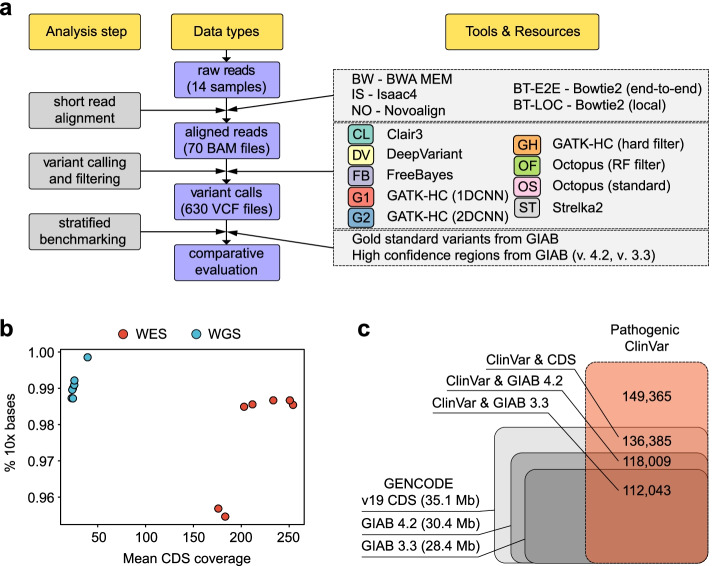
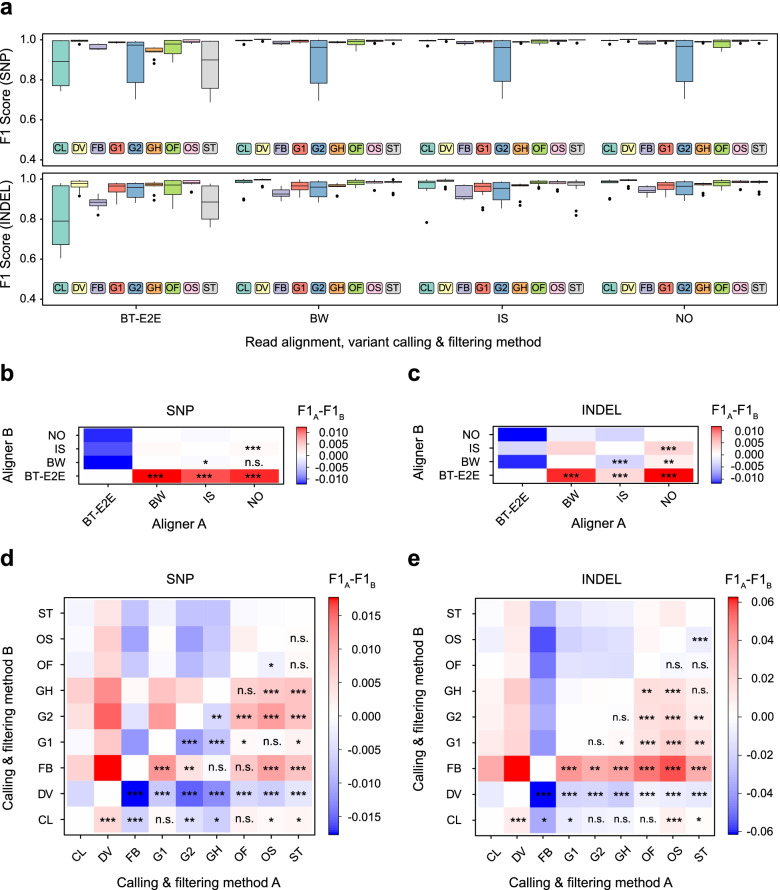
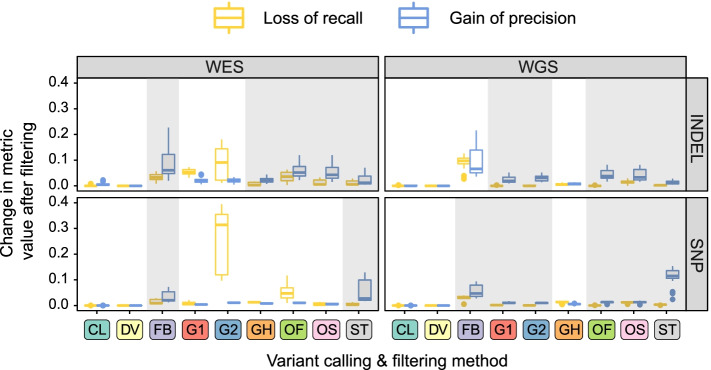
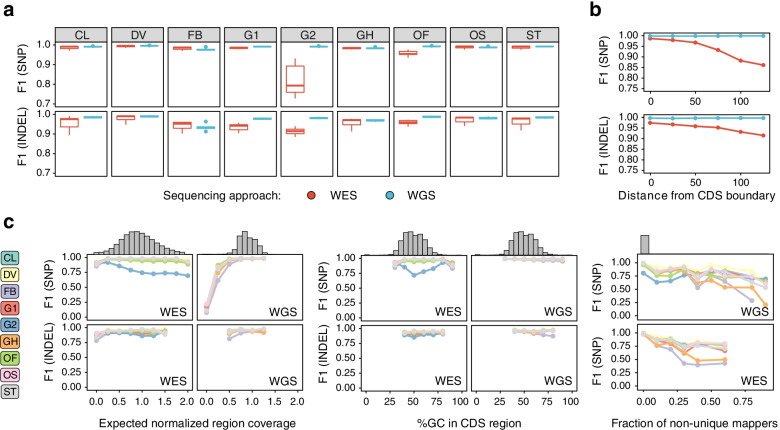
Results: In this work, we systematically evaluated the performance of 4 popular short read aligners (Bowtie2, BWA, Isaac, and Novoalign) and 9 novel and well-established variant calling and filtering methods (Clair3, DeepVariant, Octopus, GATK, FreeBayes, and Strelka2) using a set of 14 "gold standard" WES and WGS datasets available from Genome In A Bottle (GIAB) consortium. Additionally, we have indirectly evaluated each pipeline's performance using a set of 6 non-GIAB samples of African and Russian ethnicity. In our benchmark, Bowtie2 performed significantly worse than other aligners, suggesting it should not be used for medical variant calling. When other aligners were considered, the accuracy of variant discovery mostly depended on the variant caller and not the read aligner. Among the tested variant callers, DeepVariant consistently showed the best performance and the highest robustness. Other actively developed tools, such as Clair3, Octopus, and Strelka2, also performed well, although their efficiency had greater dependence on the quality and type of the input data. We have also compared the consistency of variant calls in GIAB and non-GIAB samples. With few important caveats, best-performing tools have shown little evidence of overfitting.
Conclusions: The results show surprisingly large differences in the performance of cutting-edge tools even in high confidence regions of the coding genome. This highlights the importance of regular benchmarking of quickly evolving tools and pipelines. We also discuss the need for a more diverse set of gold standard genomes that would include samples of African, Hispanic, or mixed ancestry. Additionally, there is also a need for better variant caller assessment in the repetitive regions of the coding genome.
Keywords: Benchmark; Genome in a Bottle; Performance comparison; Pipeline; Variant calling; Whole exome sequencing; Whole genome sequencing.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Karczewski KJ, Francioli LC, Tiao G, Cummings BB, Alföldi J, Wang Q, Collins RL, Laricchia KM, Ganna A, Birnbaum DP, Gauthier LD, Brand H, et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature. 2020;581:434–443. doi: 10.1038/s41586-020-2308-7. - DOI - PMC - PubMed
-
- van der Auwera GA, Carneiro MO, Hartl C, Poplin R, del Angel G, Levy-Moonshine A, Jordan T, Shakir K, Roazen D, Thibault J, Banks E, Garimella KV, et al. From FastQ data to high-confidence variant calls: the genome analysis toolkit best practices pipeline. Curr Protoc Bioinforma. 2013;10:1–10.33. - PMC - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
