

Induction of APOBEC3-mediated genomic damage in urothelium implicates BK polyomavirus (BKPyV) as a hit-and-run driver for bladder cancer
- PMID: 35194151
- PMCID: PMC8862006
- DOI: 10.1038/s41388-022-02235-8
Induction of APOBEC3-mediated genomic damage in urothelium implicates BK polyomavirus (BKPyV) as a hit-and-run driver for bladder cancer
Abstract
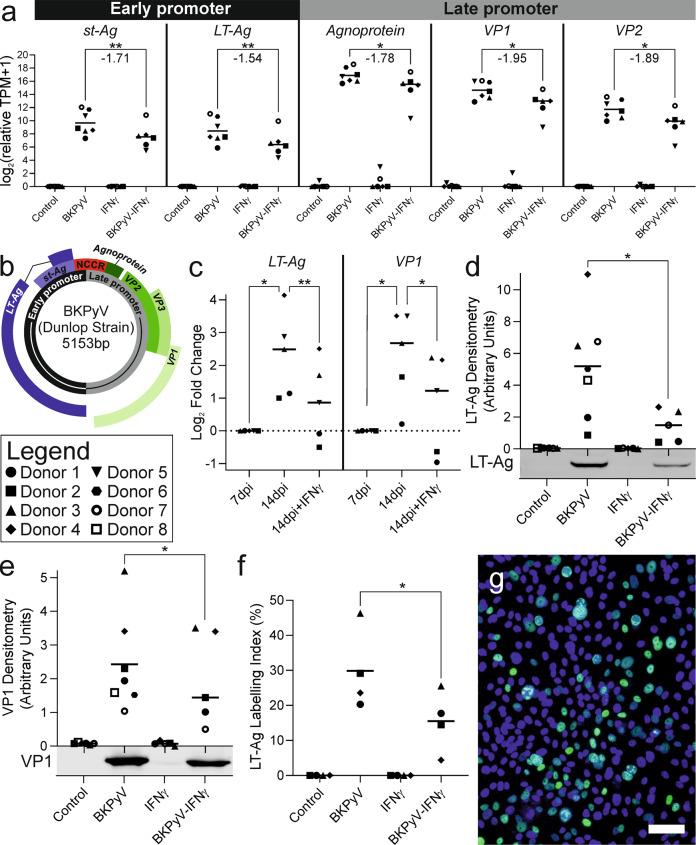
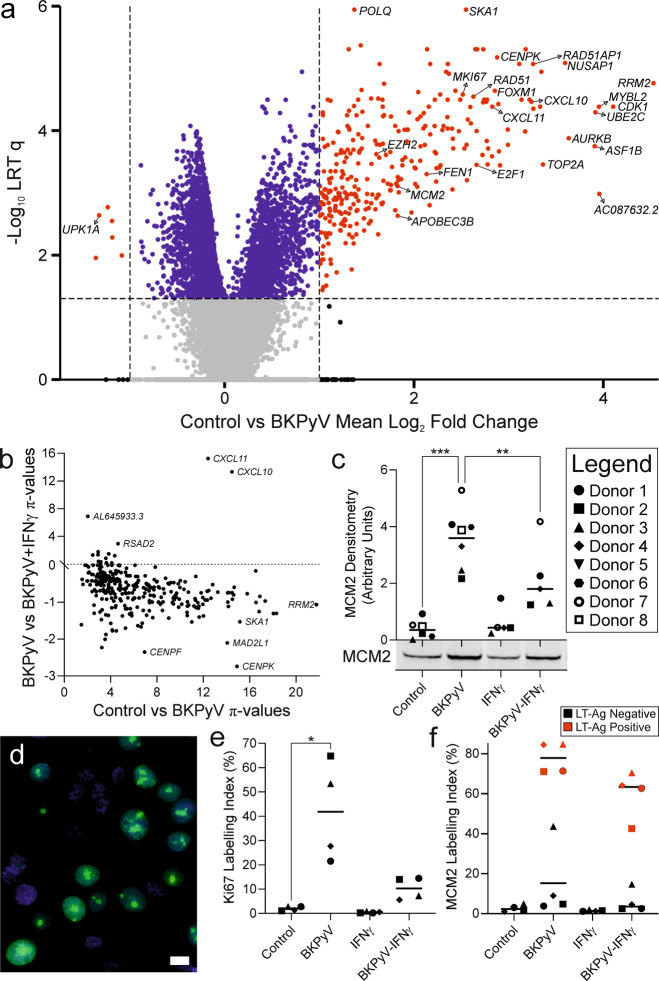
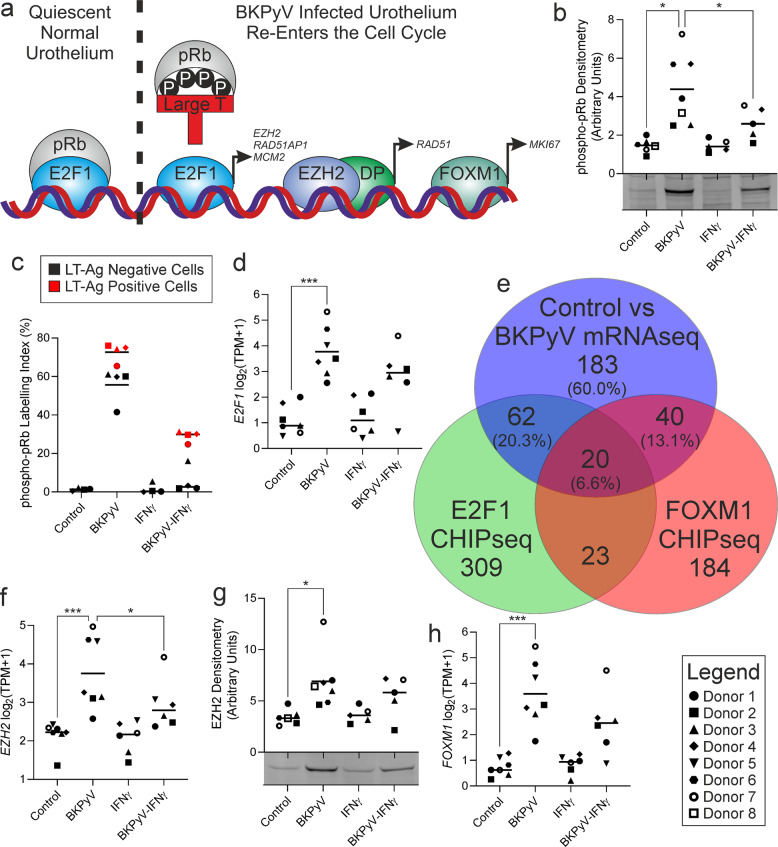
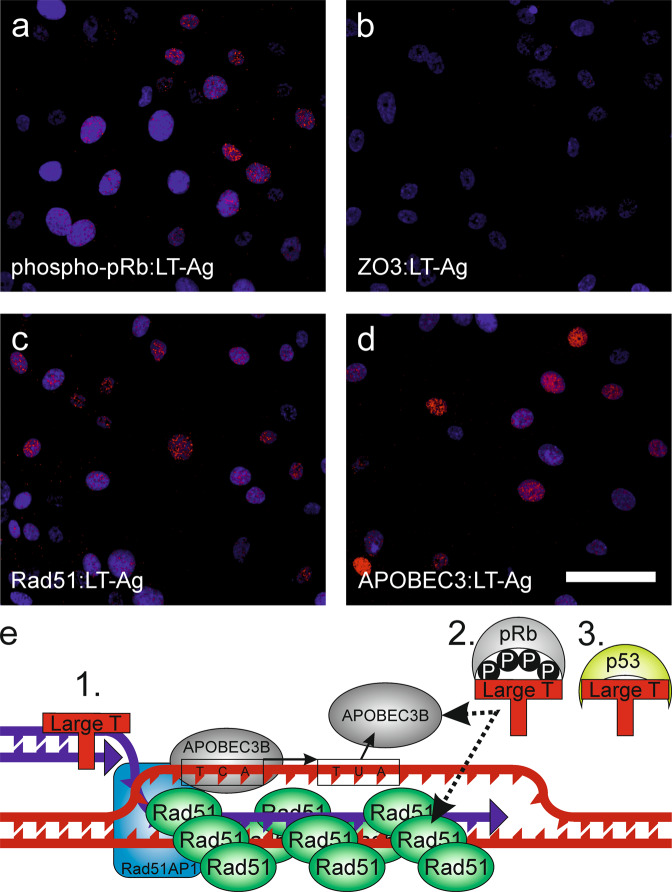
Limited understanding of bladder cancer aetiopathology hampers progress in reducing incidence. Mutational signatures show the anti-viral apolipoprotein B mRNA editing enzyme catalytic polypeptide (APOBEC) enzymes are responsible for the preponderance of mutations in bladder tumour genomes, but no causative viral agent has been identified. BK polyomavirus (BKPyV) is a common childhood infection that remains latent in the adult kidney, where reactivation leads to viruria. This study provides missing mechanistic evidence linking reactivated BKPyV-infection to bladder cancer risk. We used a mitotically-quiescent, functionally-differentiated model of normal human urothelium to examine BKPyV-infection. BKPyV-infection led to significantly elevated APOBEC3A and APOBEC3B protein, increased deaminase activity and greater numbers of apurinic/apyrimidinic sites in the host urothelial genome. BKPyV Large T antigen (LT-Ag) stimulated re-entry from G0 into the cell cycle through inhibition of retinoblastoma protein and activation of EZH2, E2F1 and FOXM1, with cells arresting in G2. The single-stranded DNA displacement loops formed in urothelial cells during BKPyV-infection interacted with LT-Ag to provide a substrate for APOBEC3-activity. Addition of interferon gamma (IFNγ) to infected urothelium suppressed expression of the viral genome. These results support reactivated BKPyV infections in adults as a risk factor for bladder cancer in immune-insufficient populations.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
