

Elastin, arterial mechanics, and stenosis
- PMID: 35196168
- PMCID: PMC9037699
- DOI: 10.1152/ajpcell.00448.2021
Elastin, arterial mechanics, and stenosis
Abstract
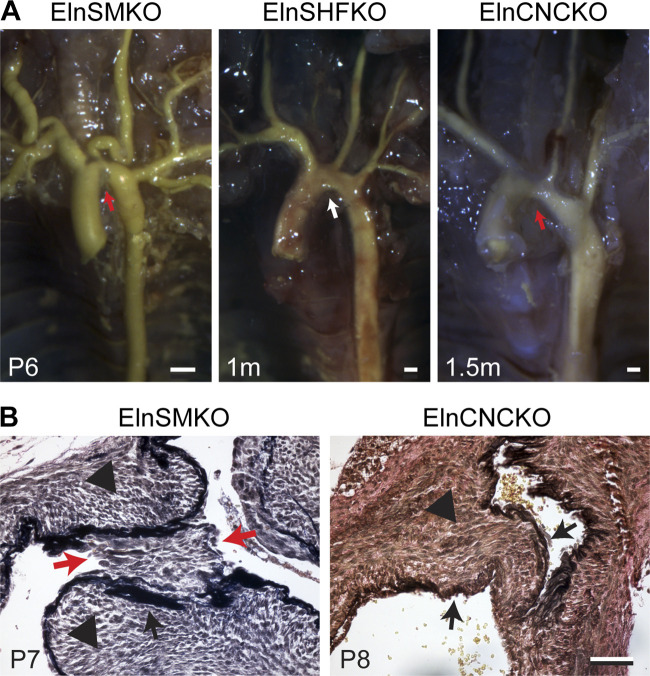
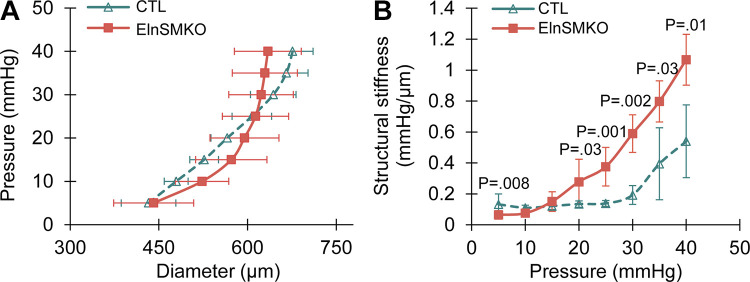
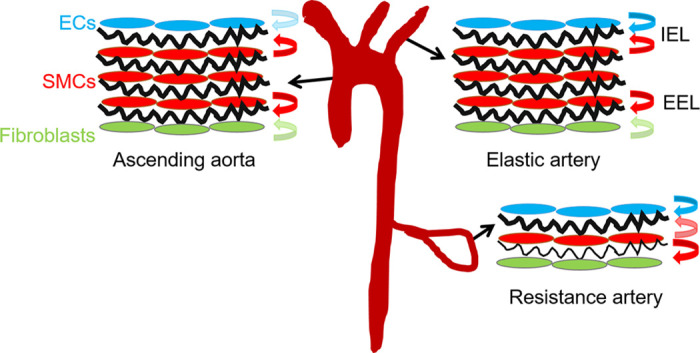
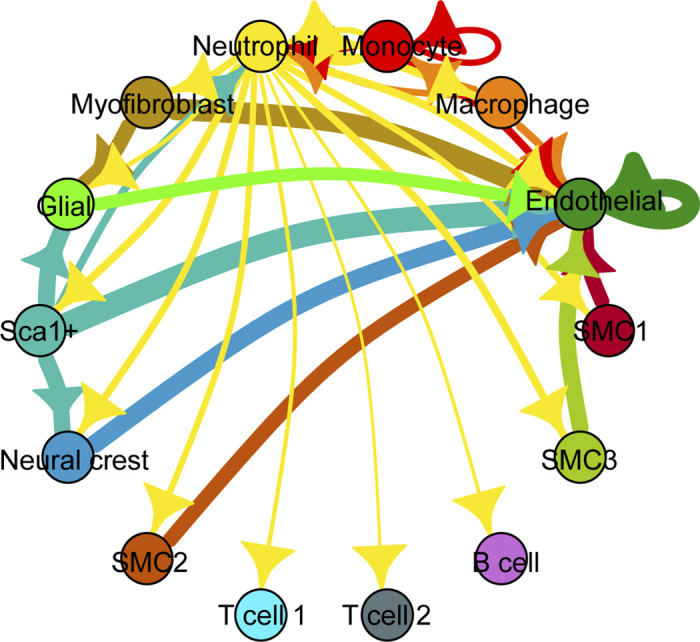
Elastin is a long-lived extracellular matrix protein that is organized into elastic fibers that provide elasticity to the arterial wall, allowing stretch and recoil with each cardiac cycle. By forming lamellar units with smooth muscle cells, elastic fibers transduce tissue-level mechanics to cell-level changes through mechanobiological signaling. Altered amounts or assembly of elastic fibers leads to changes in arterial structure and mechanical behavior that compromise cardiovascular function. In particular, genetic mutations in the elastin gene (ELN) that reduce elastin protein levels are associated with focal arterial stenosis, or narrowing of the arterial lumen, such as that seen in supravalvular aortic stenosis and Williams-Beuren syndrome. Global reduction of Eln levels in mice allows investigation of the tissue- and cell-level arterial mechanical changes and associated alterations in smooth muscle cell phenotype that may contribute to stenosis formation. A loxP-floxed Eln allele in mice highlights cell type- and developmental origin-specific mechanobiological effects of reduced elastin amounts. Eln production is required in distinct cell types for elastic layer formation in different parts of the mouse vasculature. Eln deletion in smooth muscle cells from different developmental origins in the ascending aorta leads to characteristic patterns of vascular stenosis and neointima. Dissecting the mechanobiological signaling associated with local Eln depletion and subsequent smooth muscle cell response may help develop new therapeutic interventions for elastin-related diseases.
Keywords: extracellular matrix; mechanobiology; neointima; smooth muscle cell; vascular stenosis.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures
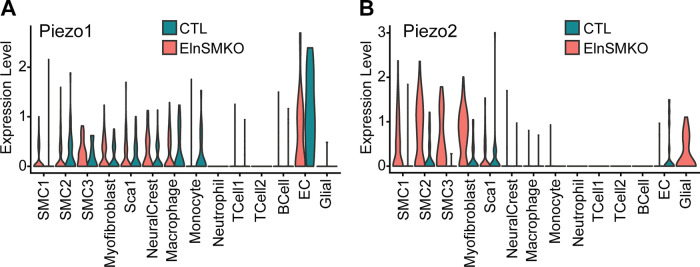
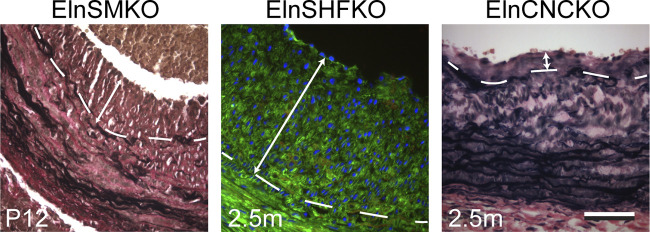
References
Publication types
MeSH terms
Substances
Grants and funding
- T32HL125241/HHS | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R56 HL152420/HL/NHLBI NIH HHS/United States
- T32HL007081/HHS | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R56HL152420/HHS | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- T32 HL125241/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources