

Abolishing the prelamin A ZMPSTE24 cleavage site leads to progeroid phenotypes with near-normal longevity in mice
- PMID: 35197292
- PMCID: PMC8892526
- DOI: 10.1073/pnas.2118695119
Abolishing the prelamin A ZMPSTE24 cleavage site leads to progeroid phenotypes with near-normal longevity in mice
Abstract
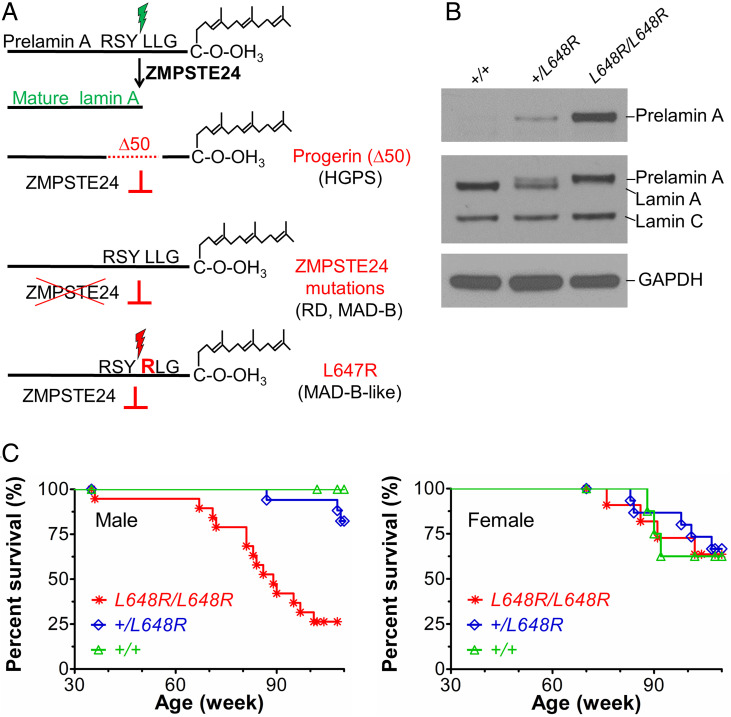
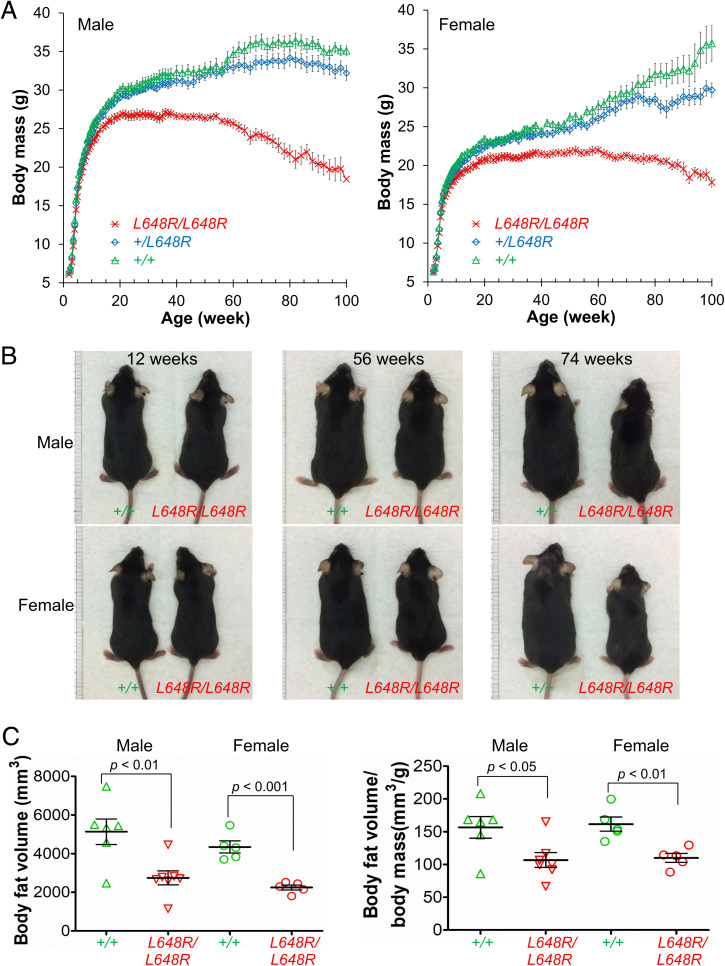
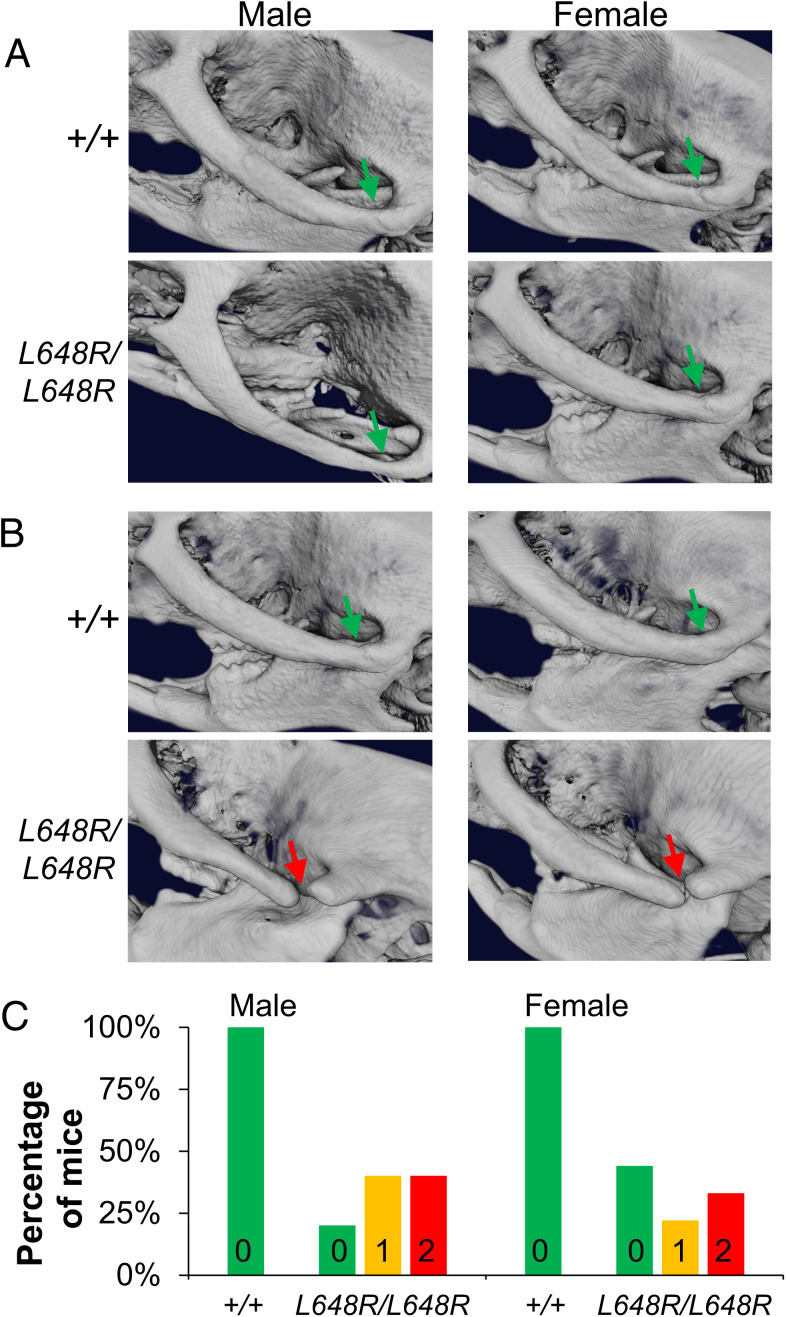
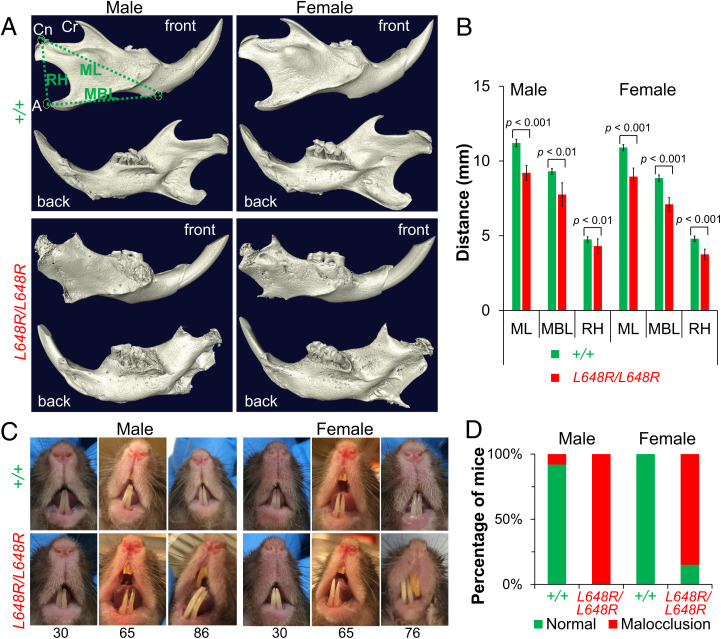
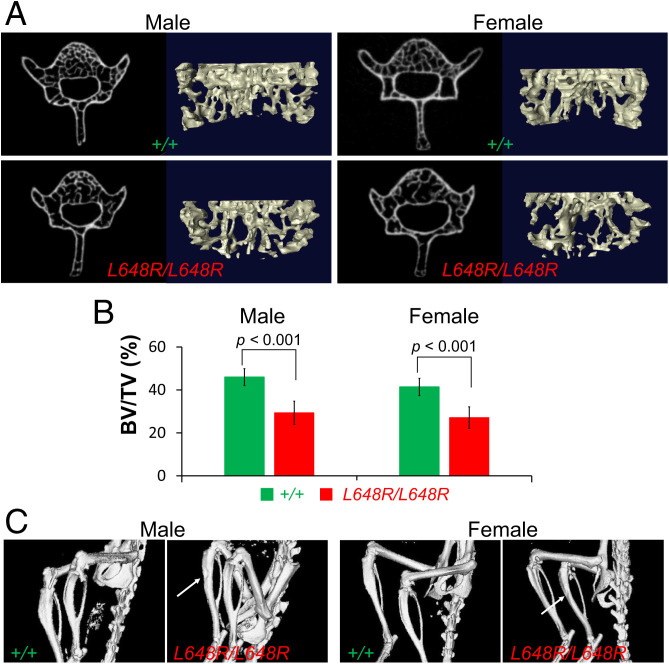
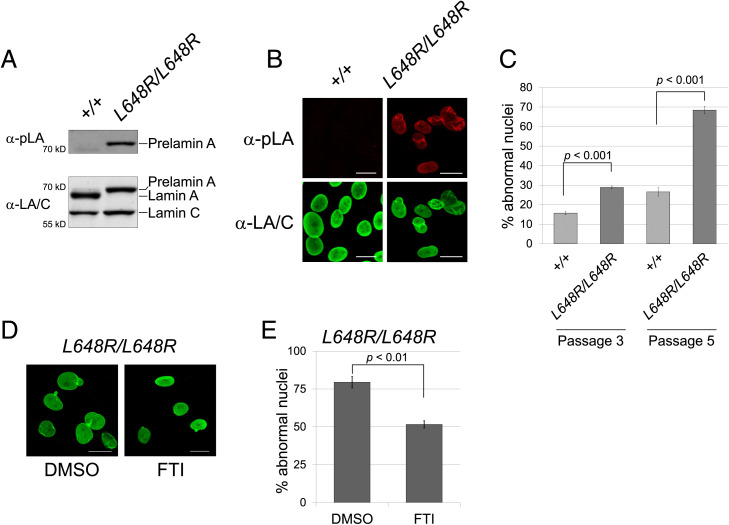
Prelamin A is a farnesylated precursor of lamin A, a nuclear lamina protein. Accumulation of the farnesylated prelamin A variant progerin, with an internal deletion including its processing site, causes Hutchinson-Gilford progeria syndrome. Loss-of-function mutations in ZMPSTE24, which encodes the prelamin A processing enzyme, lead to accumulation of full-length farnesylated prelamin A and cause related progeroid disorders. Some data suggest that prelamin A also accumulates with physiological aging. Zmpste24-/- mice die young, at ∼20 wk. Because ZMPSTE24 has functions in addition to prelamin A processing, we generated a mouse model to examine effects solely due to the presence of permanently farnesylated prelamin A. These mice have an L648R amino acid substitution in prelamin A that blocks ZMPSTE24-catalyzed processing to lamin A. The LmnaL648R/L648R mice express only prelamin and no mature protein. Notably, nearly all survive to 65 to 70 wk, with ∼40% of male and 75% of female LmnaL648R/L648R mice having near-normal lifespans of 90 wk (almost 2 y). Starting at ∼10 wk of age, LmnaL648R/L648R mice of both sexes have lower body masses than controls. By ∼20 to 30 wk of age, they exhibit detectable cranial, mandibular, and dental defects similar to those observed in Zmpste24-/- mice and have decreased vertebral bone density compared to age- and sex-matched controls. Cultured embryonic fibroblasts from LmnaL648R/L648R mice have aberrant nuclear morphology that is reversible by treatment with a protein farnesyltransferase inhibitor. These novel mice provide a model to study the effects of farnesylated prelamin A during physiological aging.
Keywords: aging; bone; lamin; nuclear envelope; progeria.
Copyright © 2022 the Author(s). Published by PNAS.
Conflict of interest statement
Competing interest statement: H.J.W. has received consulting income from Eiger BioPharmaceuticals.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
