

Early Noise-Induced Hearing Loss Accelerates Presbycusis Altering Aging Processes in the Cochlea
- PMID: 35197842
- PMCID: PMC8860087
- DOI: 10.3389/fnagi.2022.803973
Early Noise-Induced Hearing Loss Accelerates Presbycusis Altering Aging Processes in the Cochlea
Abstract
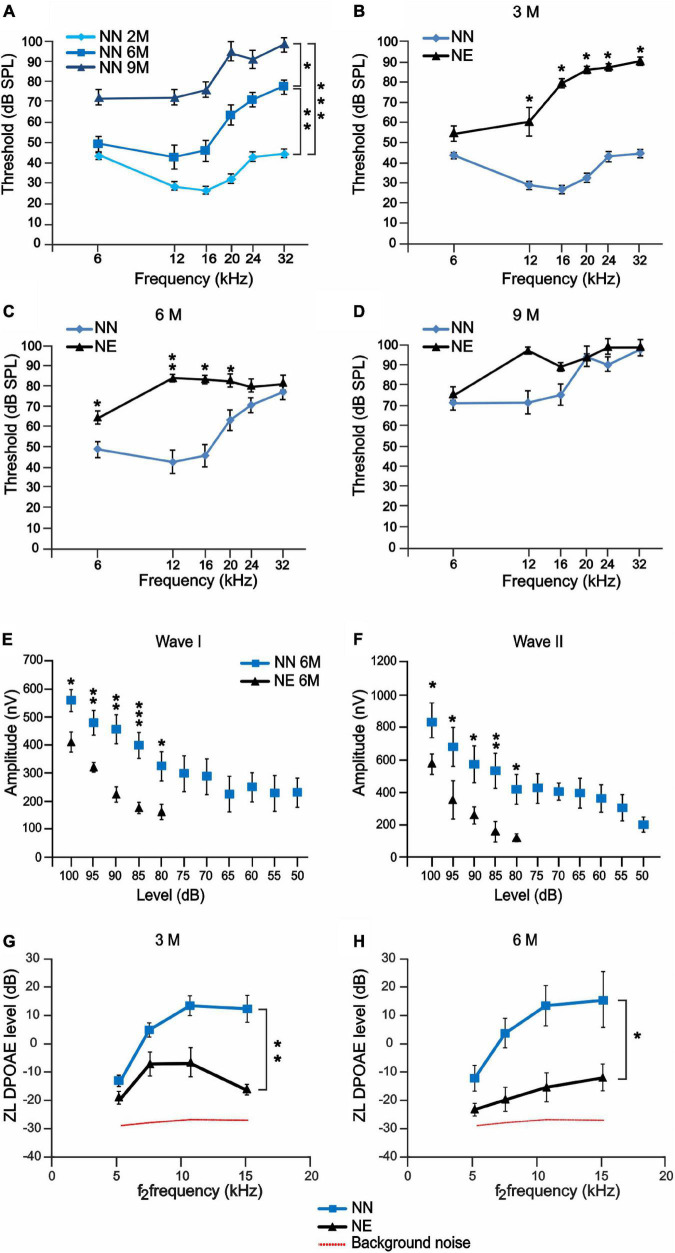
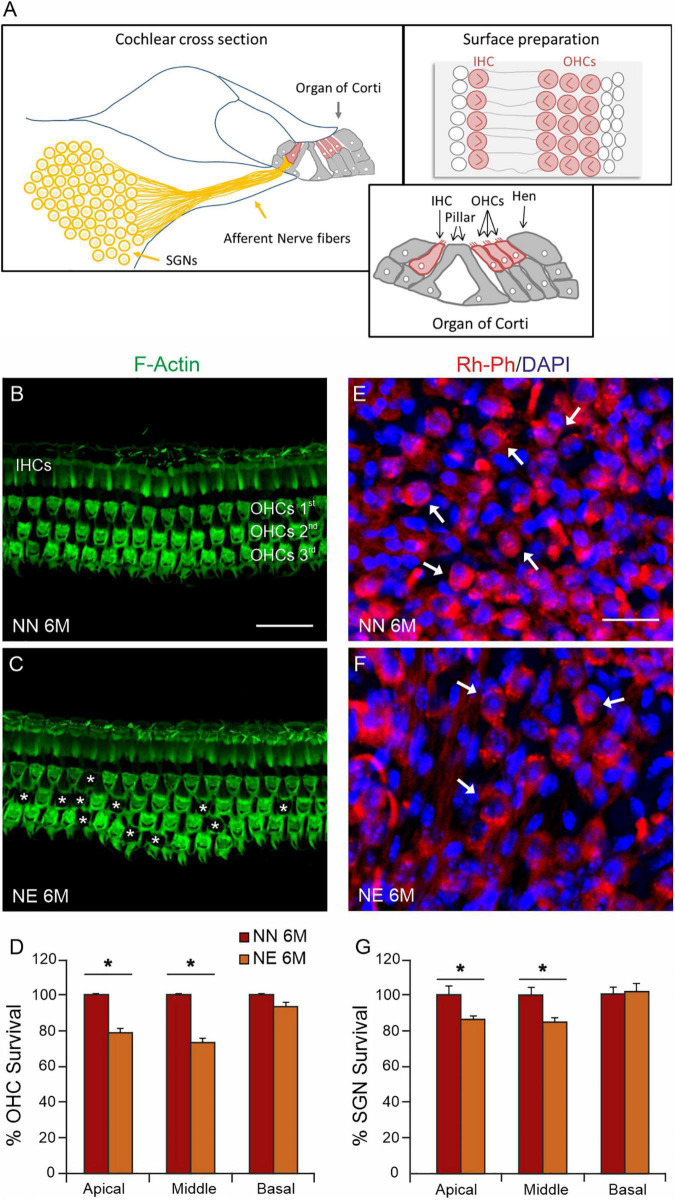
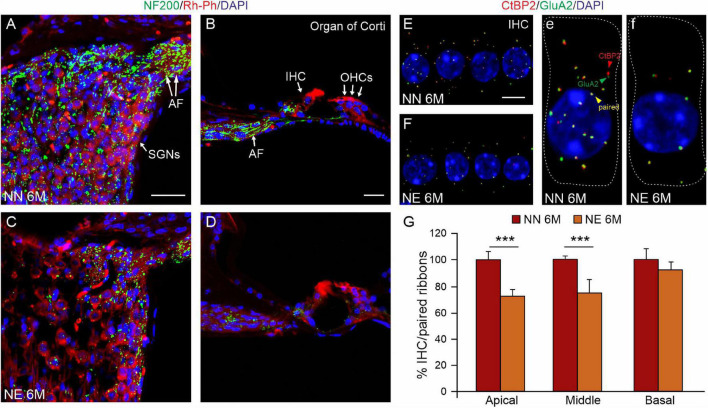
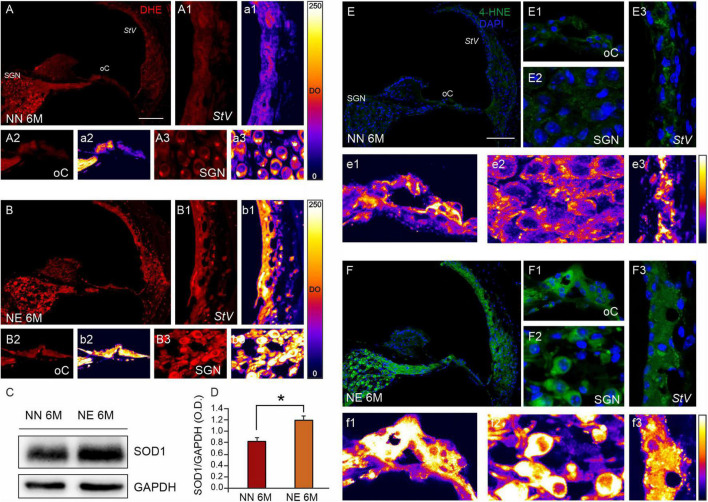
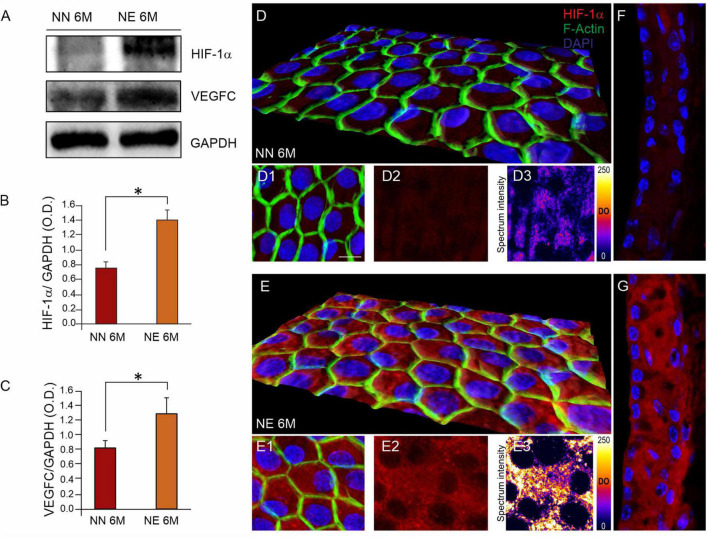
Several studies identified hearing loss as a risk factor for aging-related processes, including neurodegenerative diseases, as dementia and age-related hearing loss (ARHL). Although the association between hearing impairment in midlife and ARHL has been widely documented by epidemiological and experimental studies, the molecular mechanisms underlying this association are not fully understood. In this study, we used an established animal model of ARHL (C57BL/6 mice) to evaluate if early noise-induced hearing loss (NIHL) could affect the onset or progression of age-related cochlear dysfunction. We found that hearing loss can exacerbate ARHL, damaging sensory-neural cochlear epithelium and causing synaptopathy. Moreover, we studied common pathological markers shared between hearing loss and ARHL, demonstrating that noise exposure can worsen/accelerate redox status imbalance [increase of reactive oxygen species (ROS) production, lipid peroxidation, and dysregulation of endogenous antioxidant response] and vascular dysfunction [increased expression of hypoxia-inducible factor-1alpha (HIF-1α) and vascular endothelial growth factor C (VEGFC)] in the cochlea. Unveiling the molecular mechanisms underlying the link between hearing loss and aging processes could be valuable to identify effective therapeutic strategies to limit the effect of environmental risk factors on age-related diseases.
Keywords: acoustic trauma; age-related hearing loss; aging; hearing loss; oxidative stress; vascular dysfunction.
Copyright © 2022 Fetoni, Pisani, Rolesi, Paciello, Viziano, Moleti, Sisto, Troiani, Paludetti and Grassi.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Alvarado J. C., Fuentes-Santamaría V., Melgar-Rojas P., Valero M. L., Gabaldón-Ull M. C., Miller J. M., et al. (2015). Synergistic effects of free radical scavengers and cochlear vasodilators: a new otoprotective strategy for age-related hearing loss. Front. Aging Neurosci. 7:86. 10.3389/fnagi.2015.00086 - DOI - PMC - PubMed
-
- Bai U., Seidman M. D., Hinojosa R., Quirk W. S. (1997). Mitochondrial DNA deletions associated with aging and possibly presbycusis: a human archival temporal bone study. Am. J. Otol. 18 449–453. - PubMed
LinkOut - more resources
Full Text Sources