

Hyperexcitable arousal circuits drive sleep instability during aging
- PMID: 35201886
- PMCID: PMC9107327
- DOI: 10.1126/science.abh3021
Hyperexcitable arousal circuits drive sleep instability during aging
Abstract
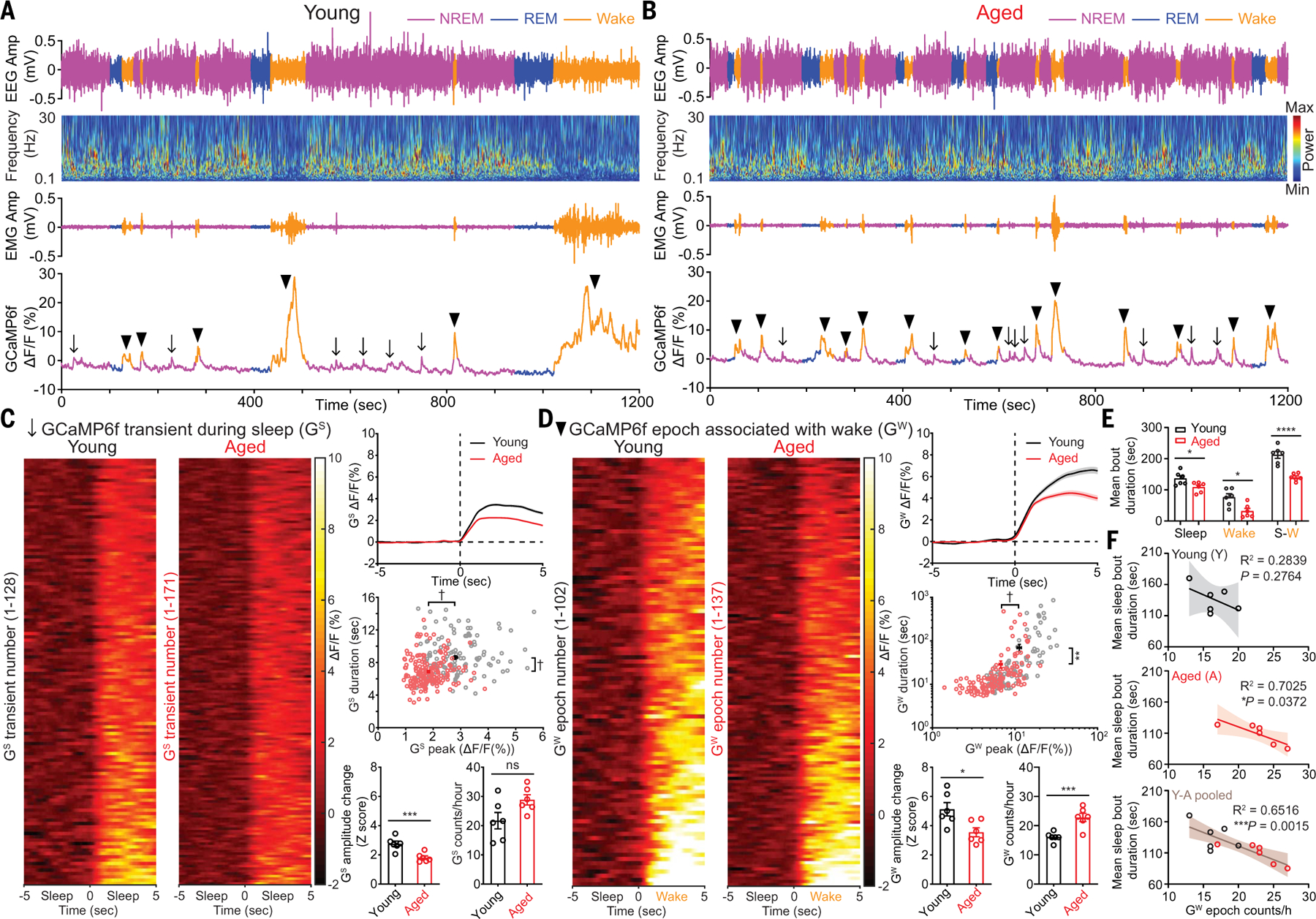
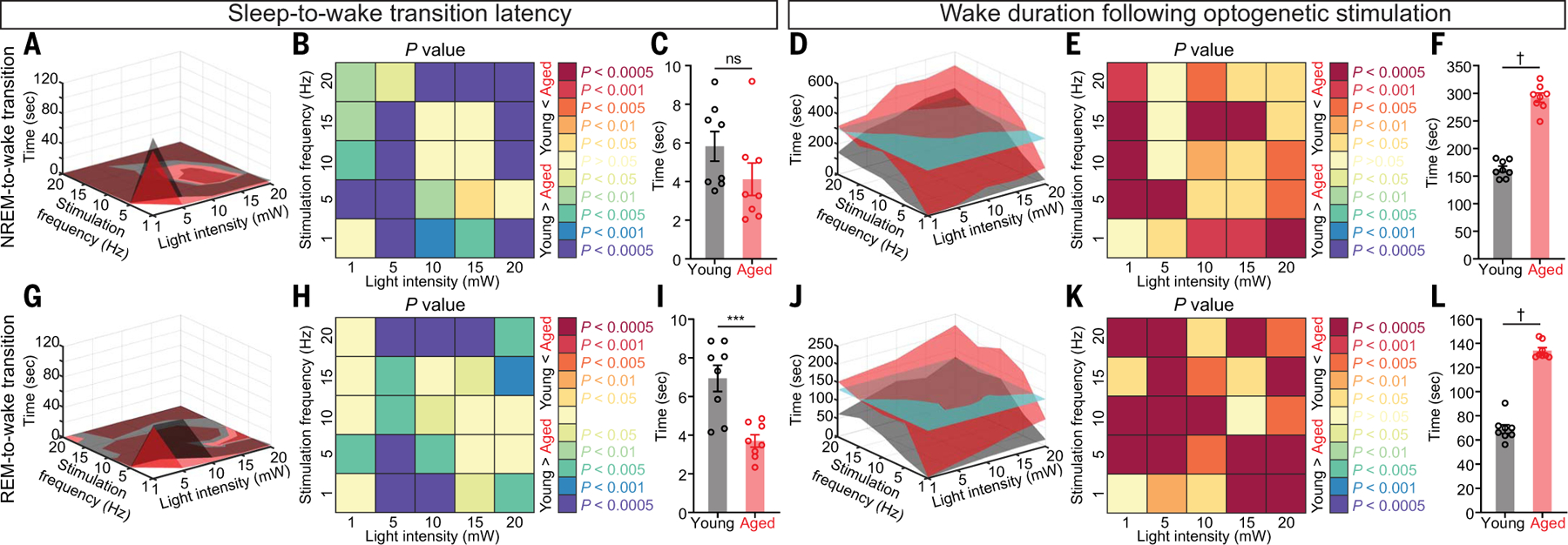
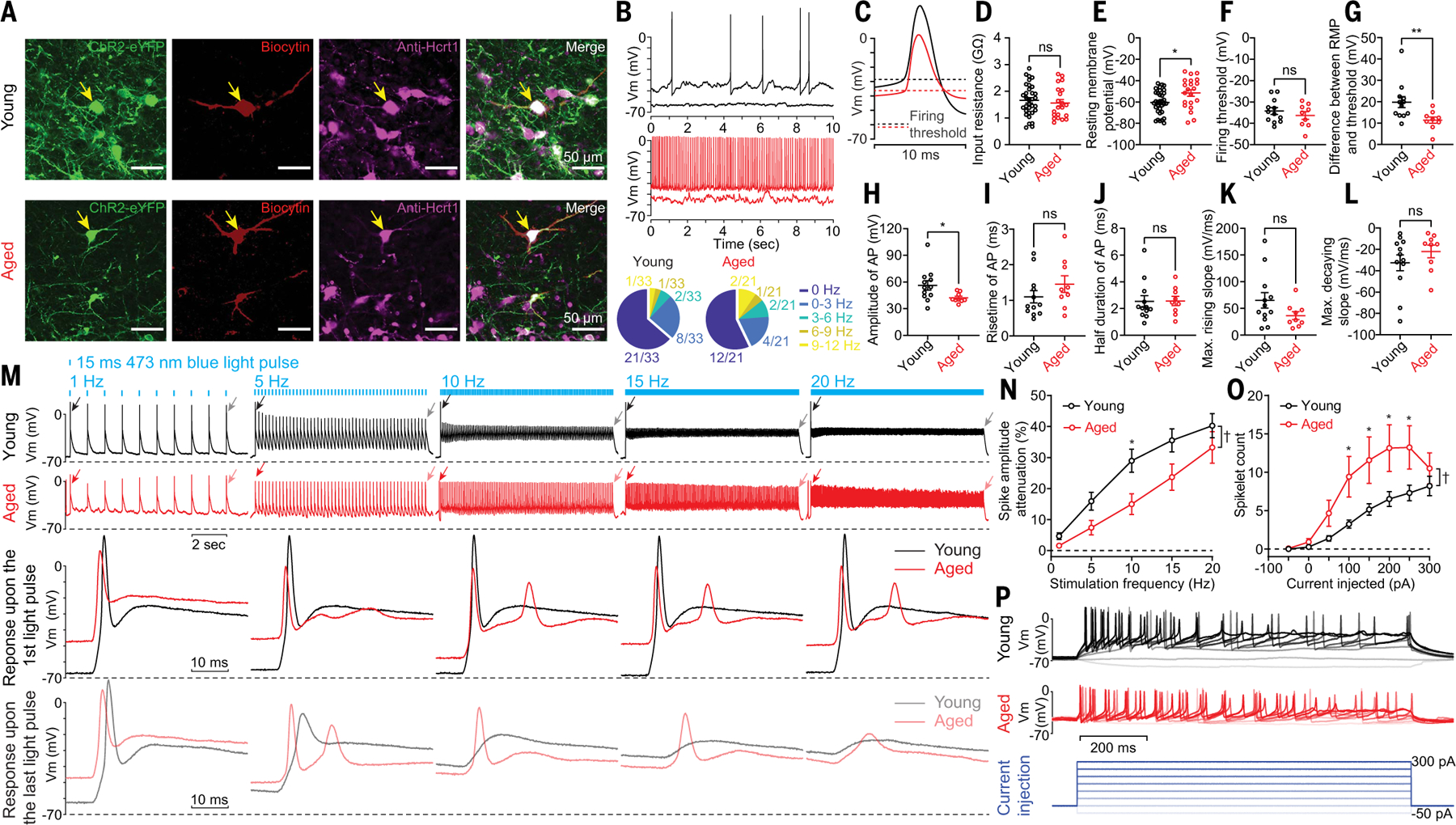
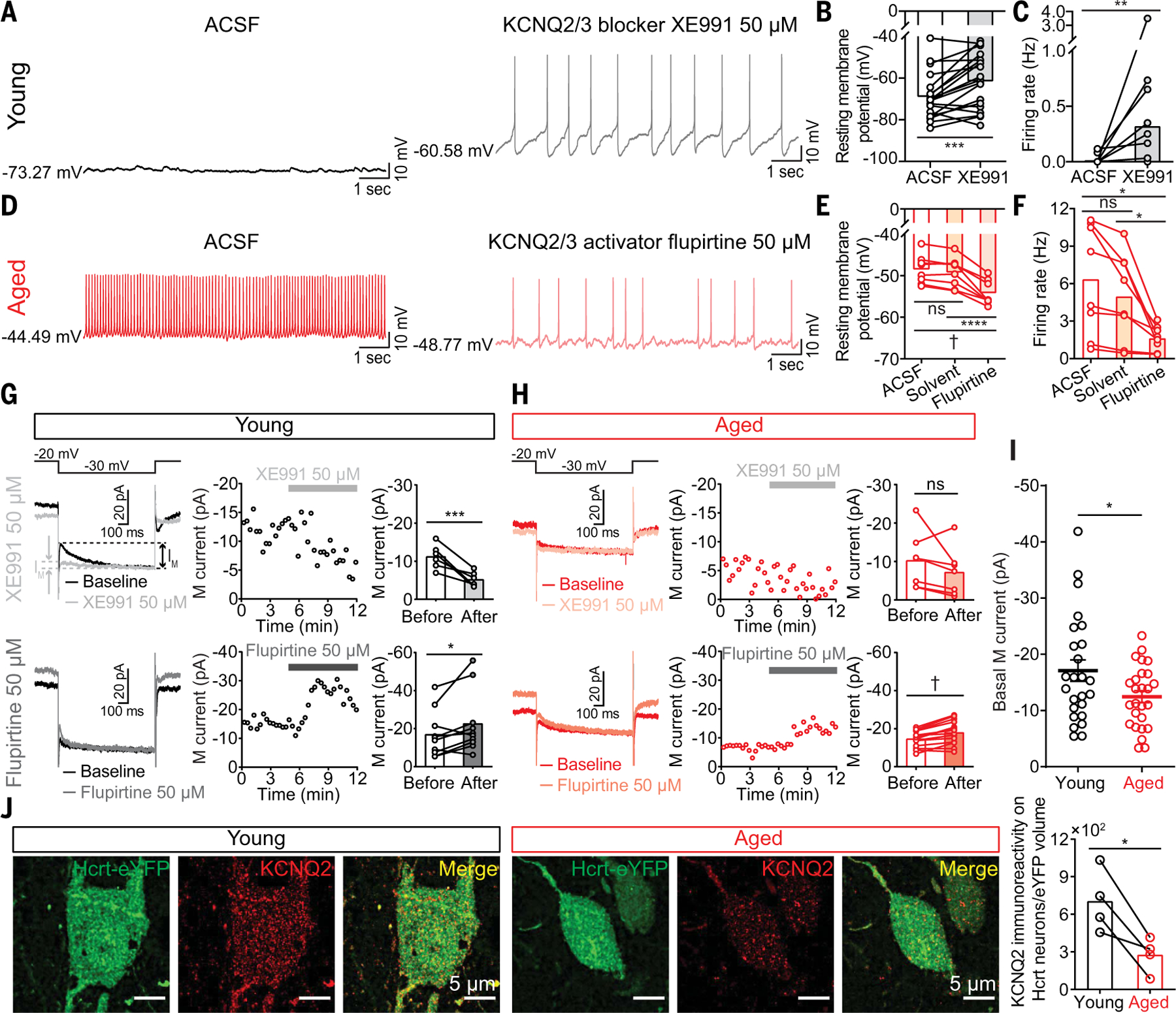
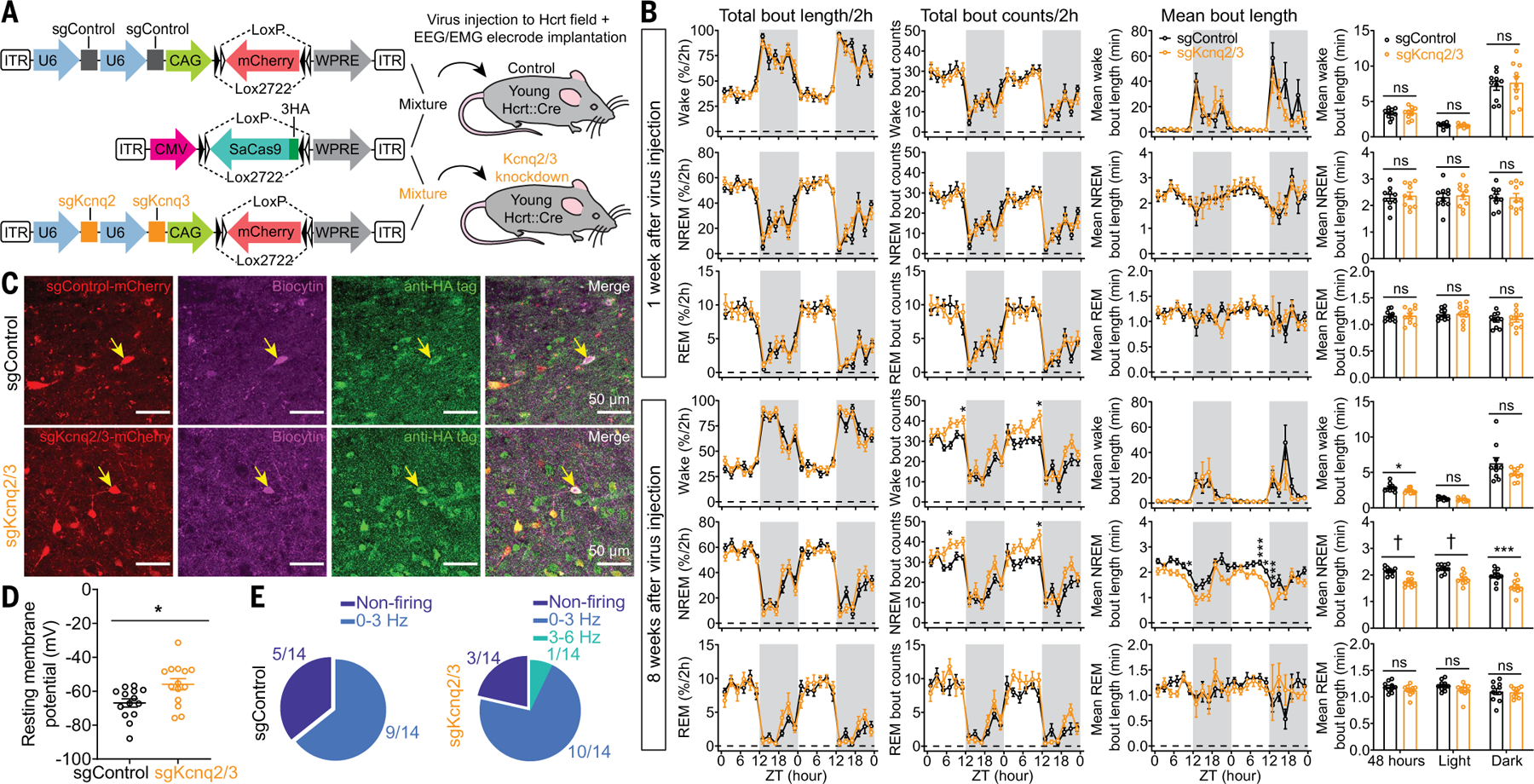
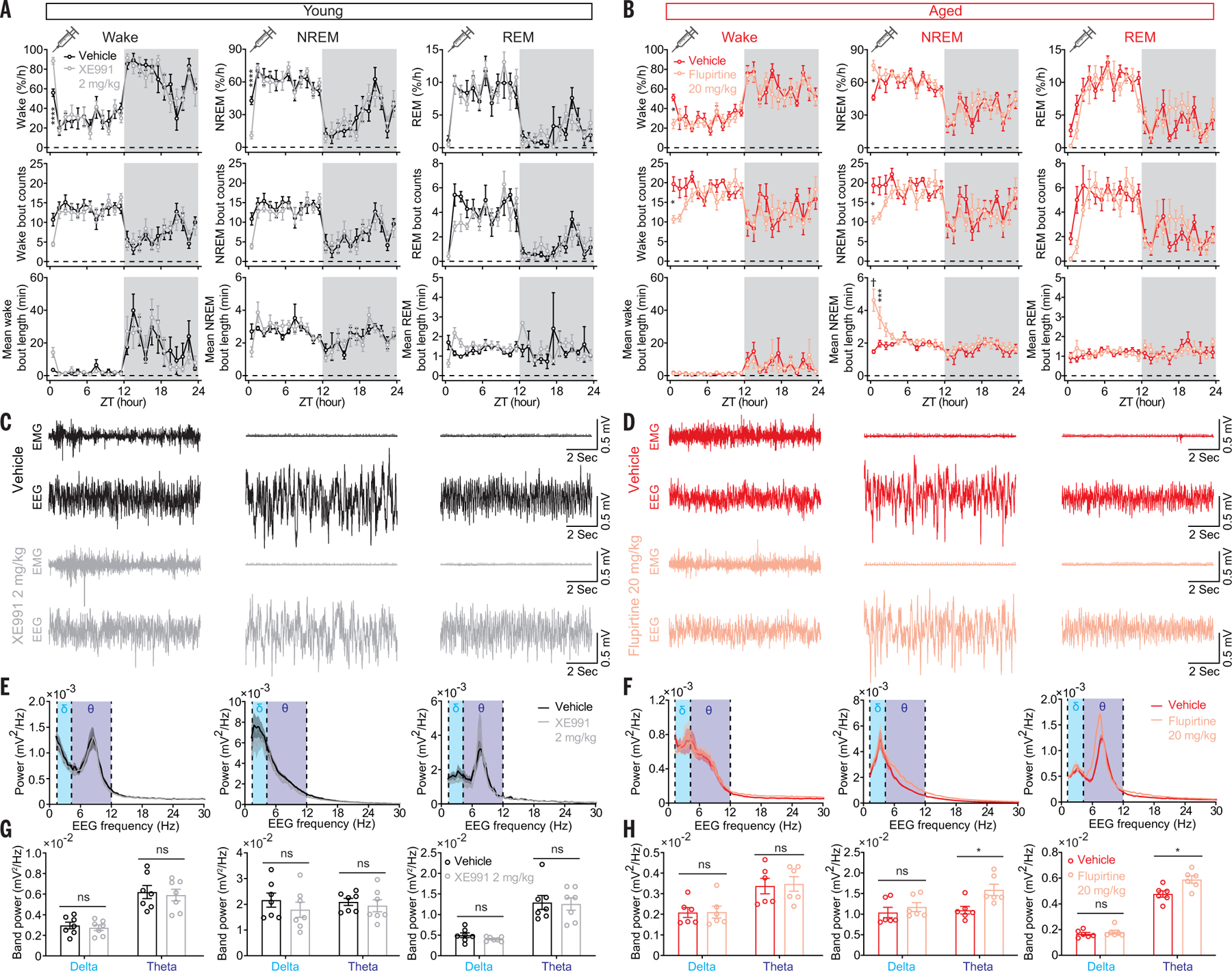
Sleep quality declines with age; however, the underlying mechanisms remain elusive. We found that hyperexcitable hypocretin/orexin (Hcrt/OX) neurons drive sleep fragmentation during aging. In aged mice, Hcrt neurons exhibited more frequent neuronal activity epochs driving wake bouts, and optogenetic activation of Hcrt neurons elicited more prolonged wakefulness. Aged Hcrt neurons showed hyperexcitability with lower KCNQ2 expression and impaired M-current, mediated by KCNQ2/3 channels. Single-nucleus RNA-sequencing revealed adaptive changes to Hcrt neuron loss in the aging brain. Disruption of Kcnq2/3 genes in Hcrt neurons of young mice destabilized sleep, mimicking aging-associated sleep fragmentation, whereas the KCNQ-selective activator flupirtine hyperpolarized Hcrt neurons and rejuvenated sleep architecture in aged mice. Our findings demonstrate a mechanism underlying sleep instability during aging and a strategy to improve sleep continuity.
Conflict of interest statement
Figures
Comment in
-
Losing sleep with age.Science. 2022 Feb 25;375(6583):816-817. doi: 10.1126/science.abo1822. Epub 2022 Feb 24. Science. 2022. PMID: 35201892
-
Too excited to sleep.Nat Rev Neurosci. 2022 May;23(5):254-255. doi: 10.1038/s41583-022-00579-4. Nat Rev Neurosci. 2022. PMID: 35288675 No abstract available.
-
Hypocretin neuron hyperexcitability in the hypothalamus: a newly discovered culprit in aging-related sleep impairment.Signal Transduct Target Ther. 2022 Jul 15;7(1):236. doi: 10.1038/s41392-022-01091-y. Signal Transduct Target Ther. 2022. PMID: 35840555 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
