

Repurposing α-Adrenoreceptor Blockers as Promising Anti-Virulence Agents in Gram-Negative Bacteria
- PMID: 35203781
- PMCID: PMC8868568
- DOI: 10.3390/antibiotics11020178
Repurposing α-Adrenoreceptor Blockers as Promising Anti-Virulence Agents in Gram-Negative Bacteria
Abstract
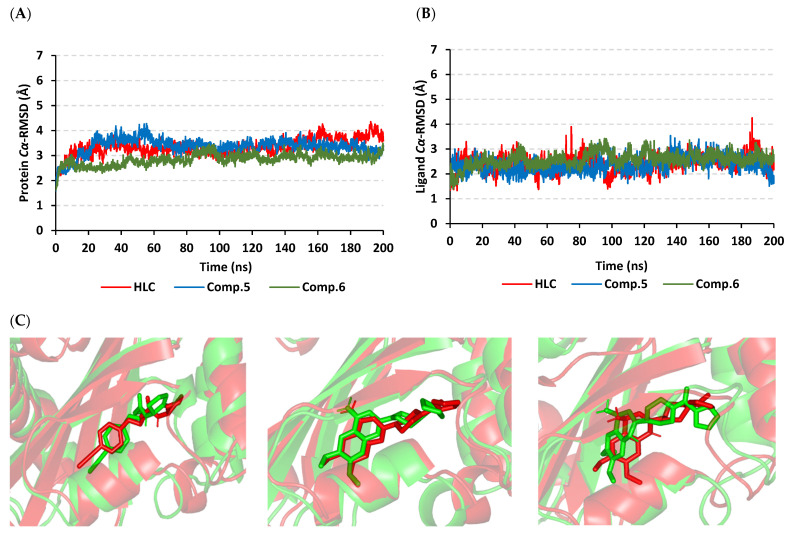
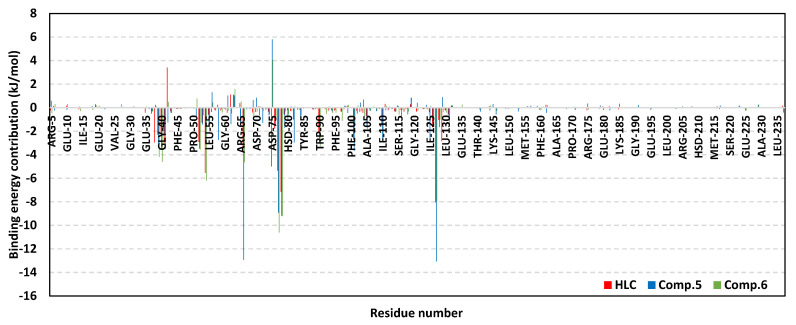
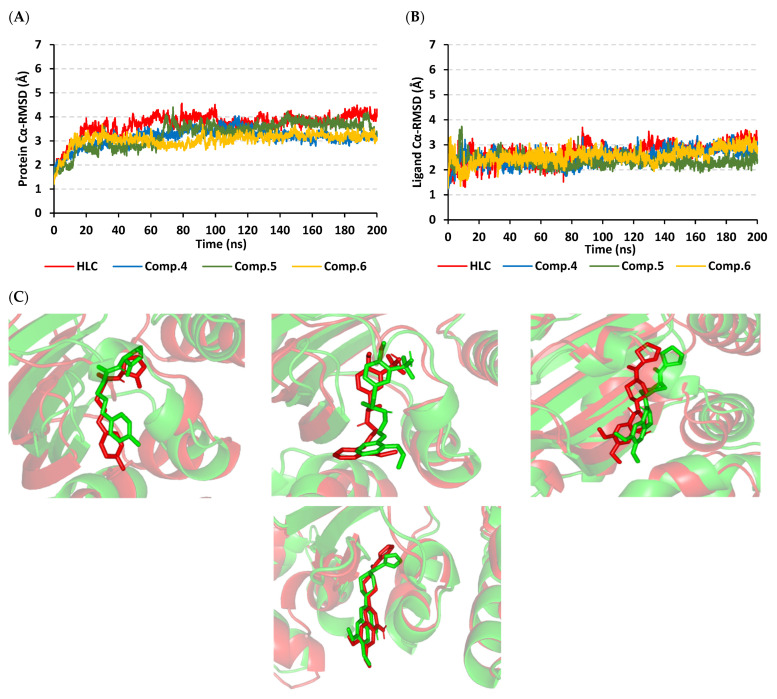
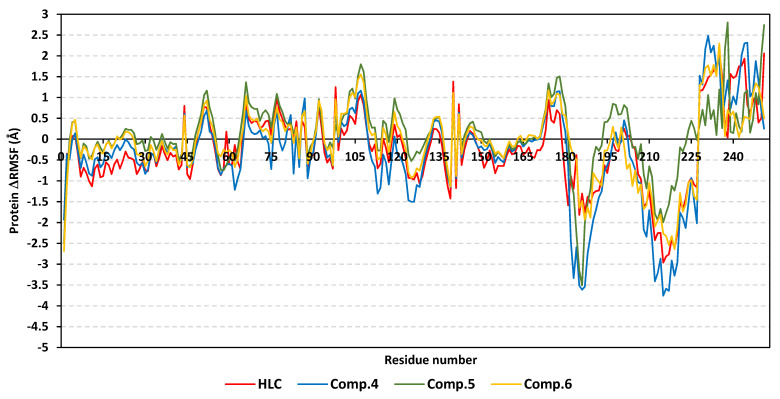
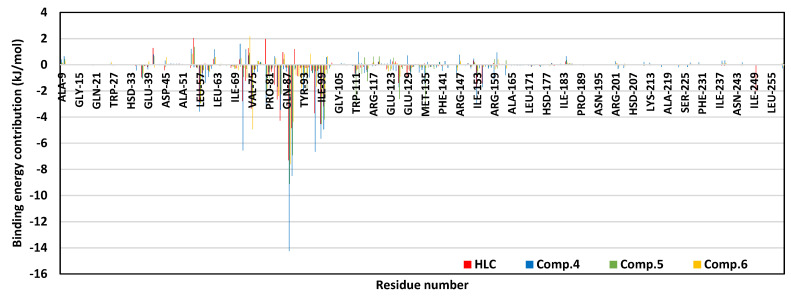
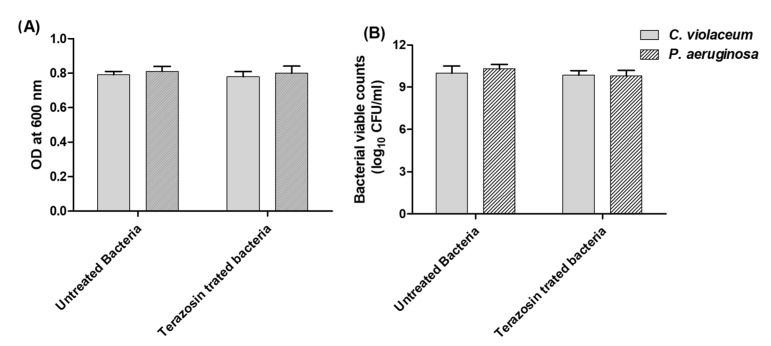
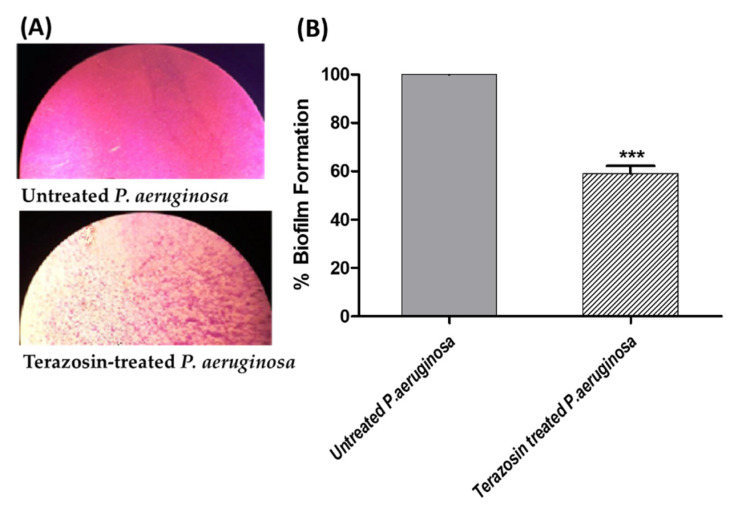
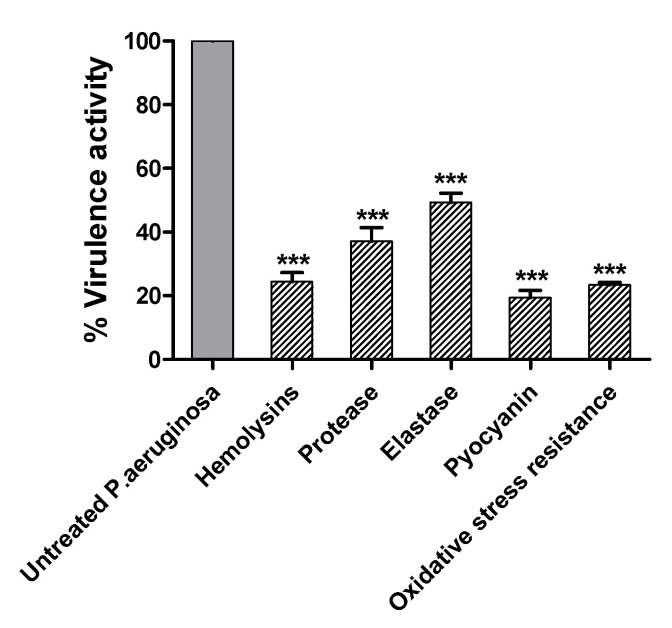
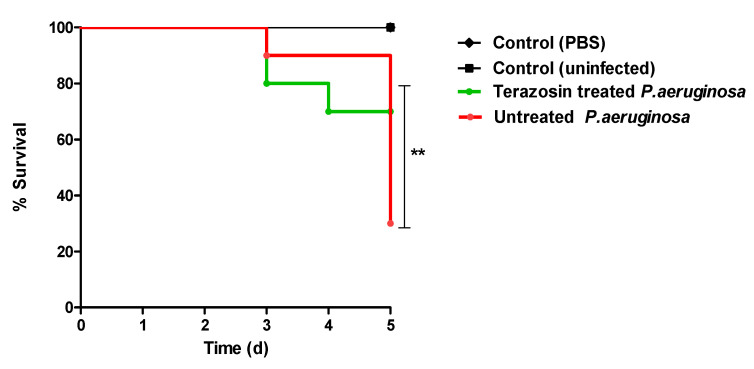
Antimicrobial resistance is among the world's most urgent public health problems. Diminishing of the virulence of bacteria is a promising approach to decrease the development of bacterial resistance. Quorum sensing (QS) systems orchestrate the bacterial virulence in inducer-receptors manner. Bacteria can spy on the cells of the host by sensing adrenergic hormones and other neurotransmitters, and in turn, these neurotransmitters can induce bacterial pathogenesis. In this direction, α-adrenergic blockers were proposed as an anti-virulence agents through inhibiting the bacterial espionage. The current study aimed to explore the α-blockers' anti-QS activities. Within comprehensive in silico investigation, the binding affinities of seven α-adrenoreceptor blockers were evaluated towards structurally different QS receptors. From the best docked α-blockers into QS receptors, terazosin was nominated to be subjected for further in vivo and in vitro anti-QS and anti-virulence activities against Chromobacterium violaceum and Pseudomonas aeruginosa. Terazosin showed a significant ability to diminish the QS-controlled pigment production in C. violaceum. Moreover, Terazosin decreased the P. aeruginosa biofilm formation and down-regulated its QS-encoding genes. Terazosin protected mice from the P. aeruginosa pathogenesis. In conclusion, α-adrenergic blockers are proposed as promising anti-virulence agents as they hinder QS receptors and inhibit bacterial espionage.
Keywords: Pseudomonas aeruginosa; bacterial virulence; quorum sensing; terazosin; α-adrenoreceptor blockers.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
Grants and funding
LinkOut - more resources
Full Text Sources
