

Nutrient Regulation of Pancreatic Islet β-Cell Secretory Capacity and Insulin Production
- PMID: 35204835
- PMCID: PMC8869698
- DOI: 10.3390/biom12020335
Nutrient Regulation of Pancreatic Islet β-Cell Secretory Capacity and Insulin Production
Abstract
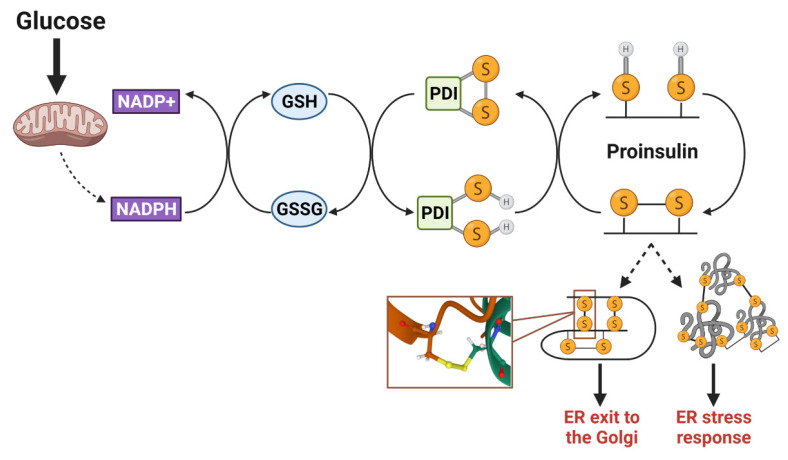
Pancreatic islet β-cells exhibit tremendous plasticity for secretory adaptations that coordinate insulin production and release with nutritional demands. This essential feature of the β-cell can allow for compensatory changes that increase secretory output to overcome insulin resistance early in Type 2 diabetes (T2D). Nutrient-stimulated increases in proinsulin biosynthesis may initiate this β-cell adaptive compensation; however, the molecular regulators of secretory expansion that accommodate the increased biosynthetic burden of packaging and producing additional insulin granules, such as enhanced ER and Golgi functions, remain poorly defined. As these adaptive mechanisms fail and T2D progresses, the β-cell succumbs to metabolic defects resulting in alterations to glucose metabolism and a decline in nutrient-regulated secretory functions, including impaired proinsulin processing and a deficit in mature insulin-containing secretory granules. In this review, we will discuss how the adaptative plasticity of the pancreatic islet β-cell's secretory program allows insulin production to be carefully matched with nutrient availability and peripheral cues for insulin signaling. Furthermore, we will highlight potential defects in the secretory pathway that limit or delay insulin granule biosynthesis, which may contribute to the decline in β-cell function during the pathogenesis of T2D.
Keywords: ER function; Golgi; beta-cell function; glutathione; granule trafficking; insulin granule; insulin secretion; proinsulin; secretory granule biogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Baker E., Blundell T.L., Cutfield J.F., Cutfield S.M., Dodson E.J., Dodson G.G., Hodgkin D.M.C., Hubbard R.E., Isaacs N.W., Reynolds C.D., et al. The structure of 2Zn pig insulin crystals at 1.5 Å resolution. Philos. Trans. R. Soc. B Biol. Sci. 1988;319:369–456. doi: 10.1098/rstb.1988.0058. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
