

Cupriavidus metallidurans CH34 Possesses Aromatic Catabolic Versatility and Degrades Benzene in the Presence of Mercury and Cadmium
- PMID: 35208938
- PMCID: PMC8879955
- DOI: 10.3390/microorganisms10020484
Cupriavidus metallidurans CH34 Possesses Aromatic Catabolic Versatility and Degrades Benzene in the Presence of Mercury and Cadmium
Abstract
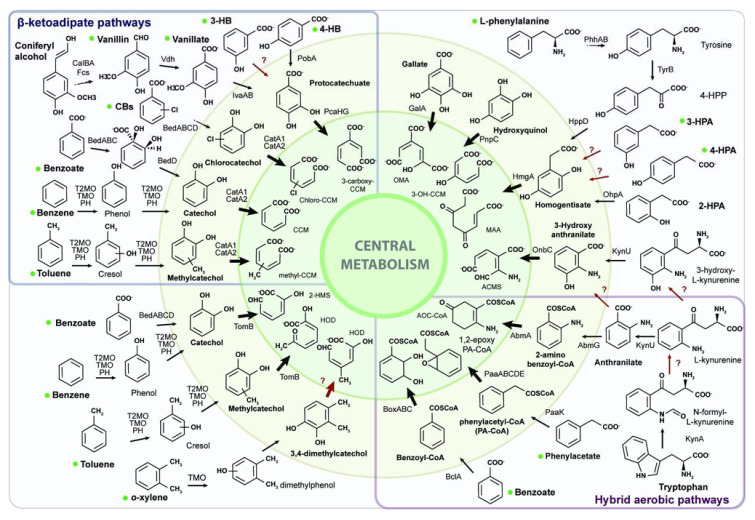
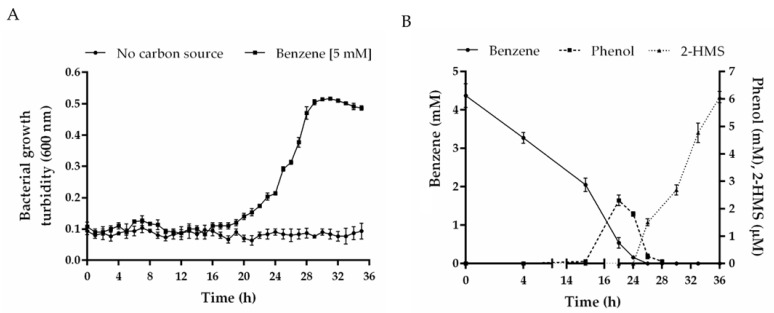
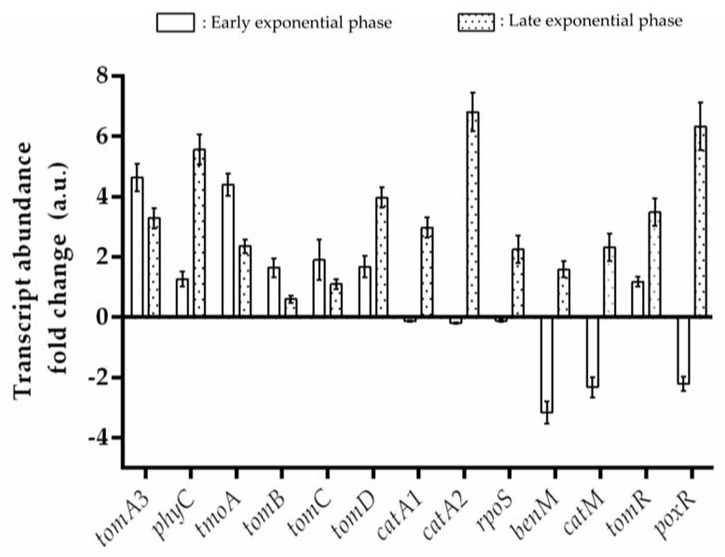
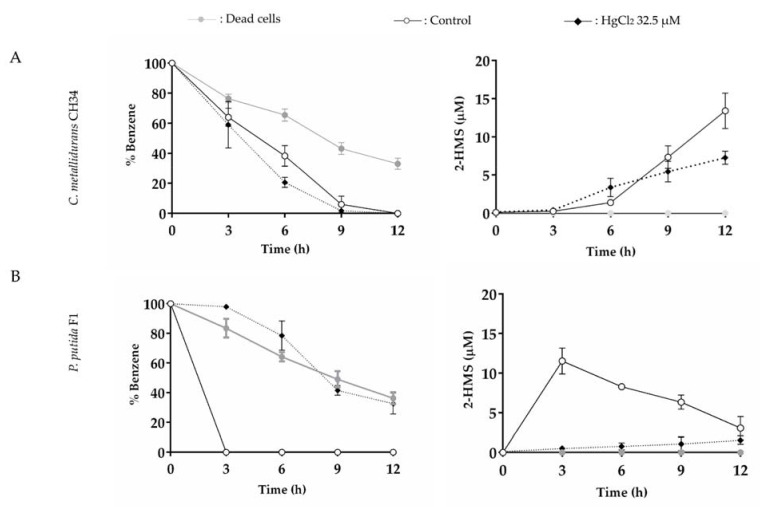
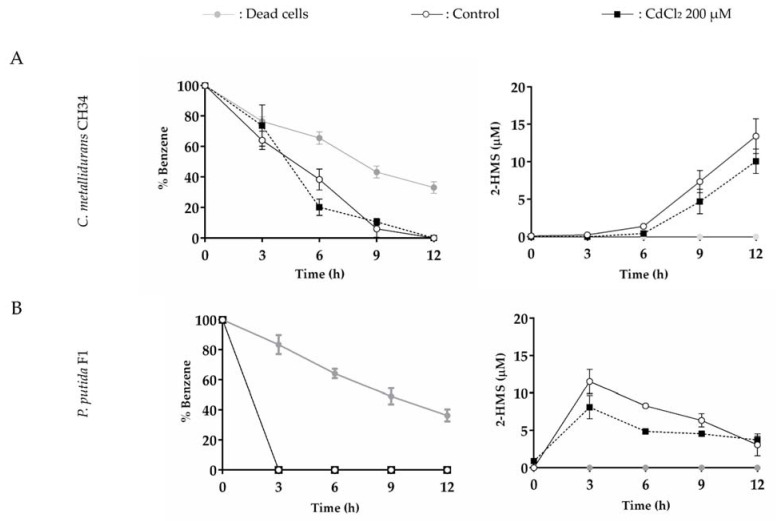
Heavy metal co-contamination in crude oil-polluted environments may inhibit microbial bioremediation of hydrocarbons. The model heavy metal-resistant bacterium Cupriavidus metallidurans CH34 possesses cadmium and mercury resistance, as well as genes related to the catabolism of hazardous BTEX aromatic hydrocarbons. The aims of this study were to analyze the aromatic catabolic potential of C. metallidurans CH34 and to determine the functionality of the predicted benzene catabolic pathway and the influence of cadmium and mercury on benzene degradation. Three chromosome-encoded bacterial multicomponent monooxygenases (BMMs) are involved in benzene catabolic pathways. Growth assessment, intermediates identification, and gene expression analysis indicate the functionality of the benzene catabolic pathway. Strain CH34 degraded benzene via phenol and 2-hydroxymuconic semialdehyde. Transcriptional analyses revealed a transition from the expression of catechol 2,3-dioxygenase (tomB) in the early exponential phase to catechol 1,2-dioxygenase (catA1 and catA2) in the late exponential phase. The minimum inhibitory concentration to Hg (II) and Cd (II) was significantly lower in the presence of benzene, demonstrating the effect of co-contamination on bacterial growth. Notably, this study showed that C. metallidurans CH34 degraded benzene in the presence of Hg (II) or Cd (II).
Keywords: Cupriavidus metallidurans; aromatic catabolism; bacterial multicomponent monooxygenase; benzene; cadmium; mercury.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures


References
-
- Liu S.H., Lai C.Y., Ye J.W., Lin C.W. Increasing removal of benzene from groundwater using stacked tubular air-cathode microbial fuel cells. J. Clean. Prod. 2018;194:78–84. doi: 10.1016/j.jclepro.2018.05.138. - DOI
-
- van der Perk M. Soil and Water Contamination. 2nd ed. CRC Press; Leiden, The Netherland: 2014. p. 167. Organic Pollutants.
-
- Nojiri H., Tsuda M., Kamagata Y., Fukuda M. Biodegradative Bacteria: How Bacteria Degrade, Survive, Adapt, and Evolve. Springer; Tokyo, Japan: 2014. p. 228. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
