

MiR-30 promotes fatty acid beta-oxidation and endothelial cell dysfunction and is a circulating biomarker of coronary microvascular dysfunction in pre-clinical models of diabetes
- PMID: 35209901
- PMCID: PMC8876371
- DOI: 10.1186/s12933-022-01458-z
MiR-30 promotes fatty acid beta-oxidation and endothelial cell dysfunction and is a circulating biomarker of coronary microvascular dysfunction in pre-clinical models of diabetes
Abstract
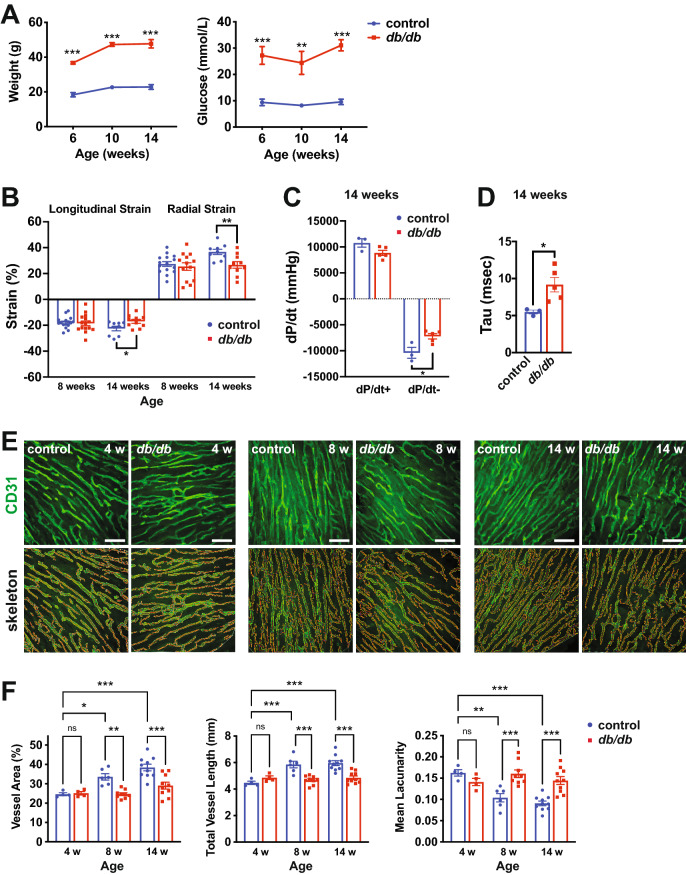
Background: Type 2 diabetes (T2D) is associated with coronary microvascular dysfunction, which is thought to contribute to compromised diastolic function, ultimately culminating in heart failure with preserved ejection fraction (HFpEF). The molecular mechanisms remain incompletely understood, and no early diagnostics are available. We sought to gain insight into biomarkers and potential mechanisms of microvascular dysfunction in obese mouse (db/db) and lean rat (Goto-Kakizaki) pre-clinical models of T2D-associated diastolic dysfunction.
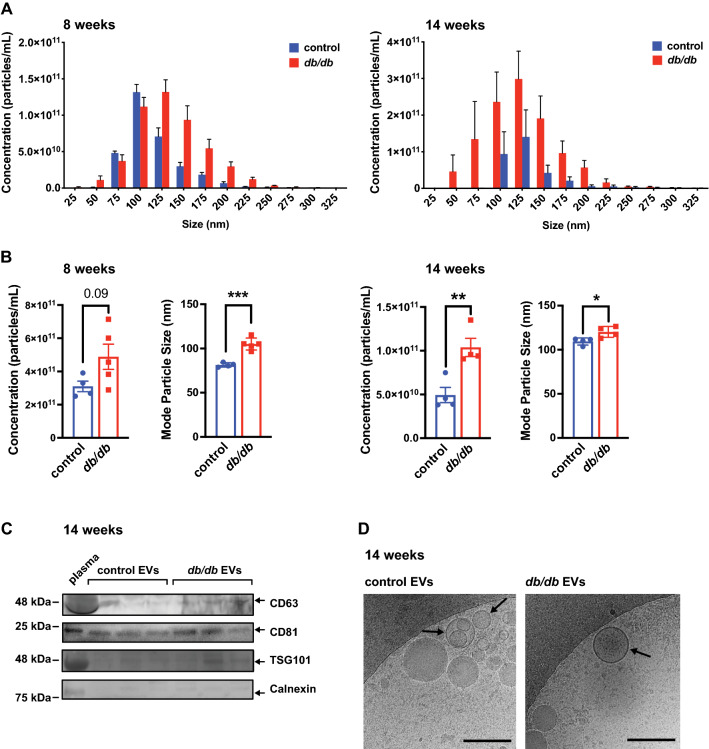
Methods: The microRNA (miRNA) content of circulating extracellular vesicles (EVs) was assessed in T2D models to identify biomarkers of coronary microvascular dysfunction/rarefaction. The potential source of circulating EV-encapsulated miRNAs was determined, and the mechanisms of induction and the function of candidate miRNAs were assessed in endothelial cells (ECs).
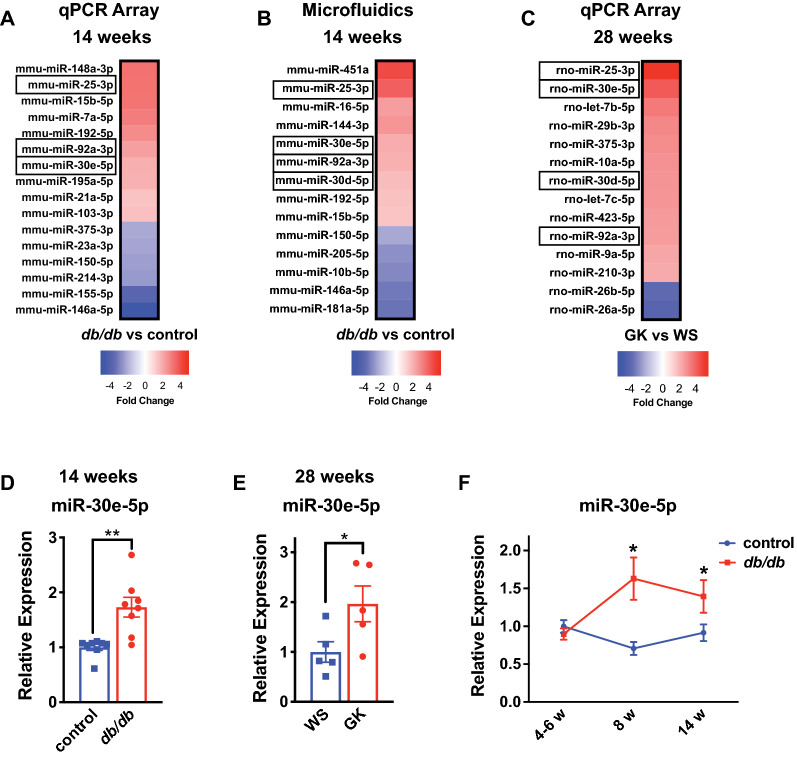
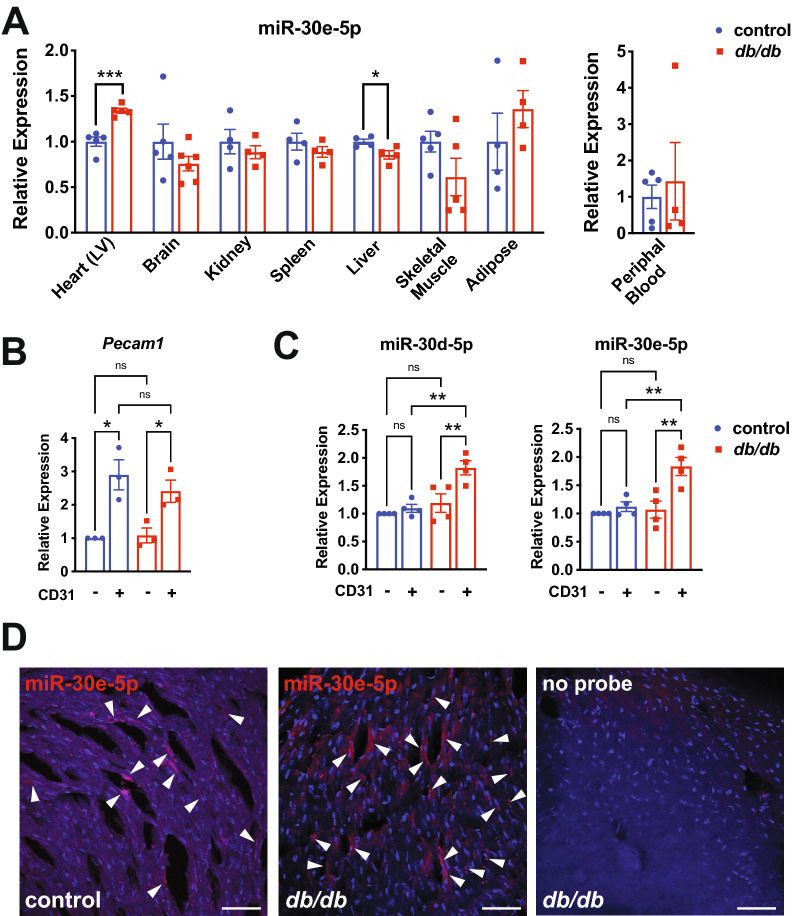
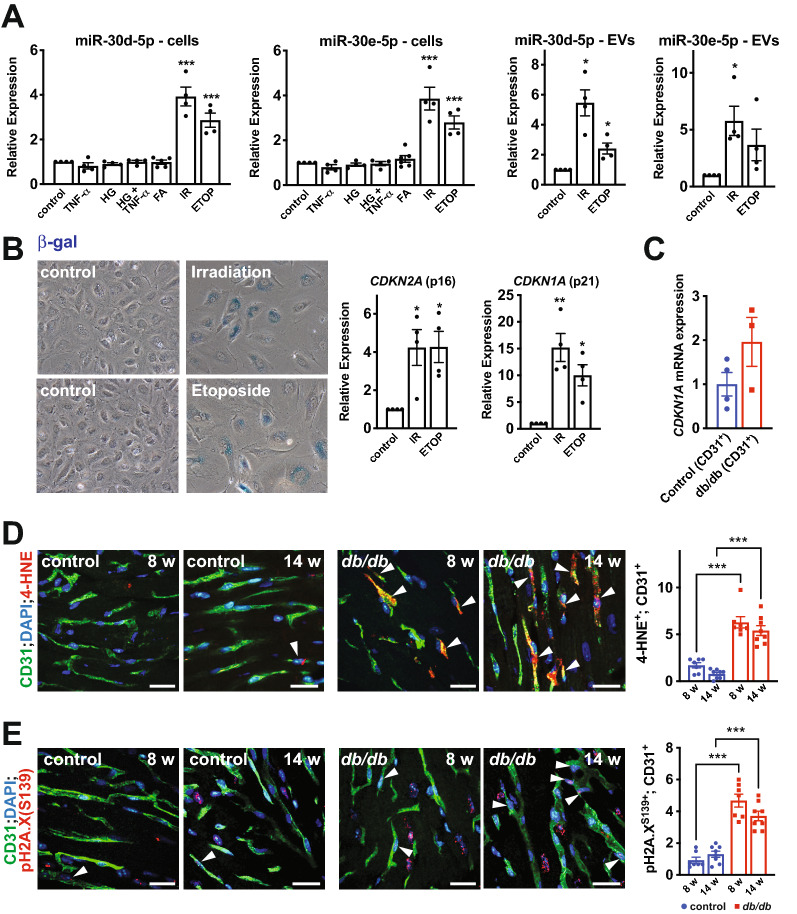
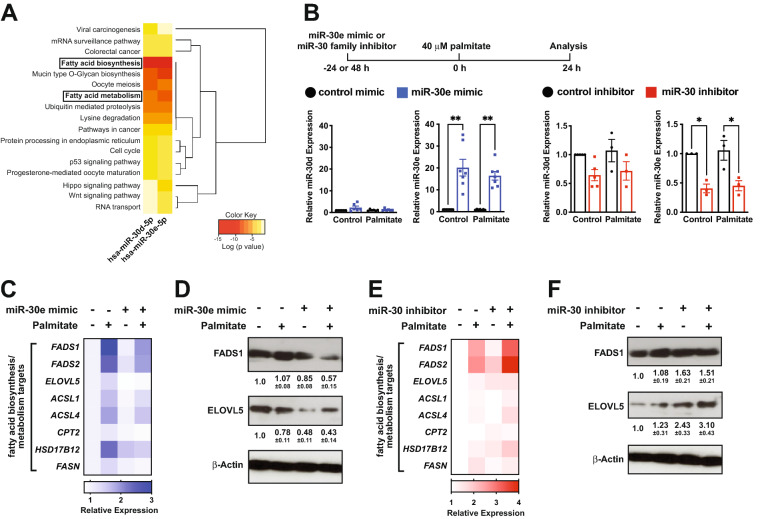
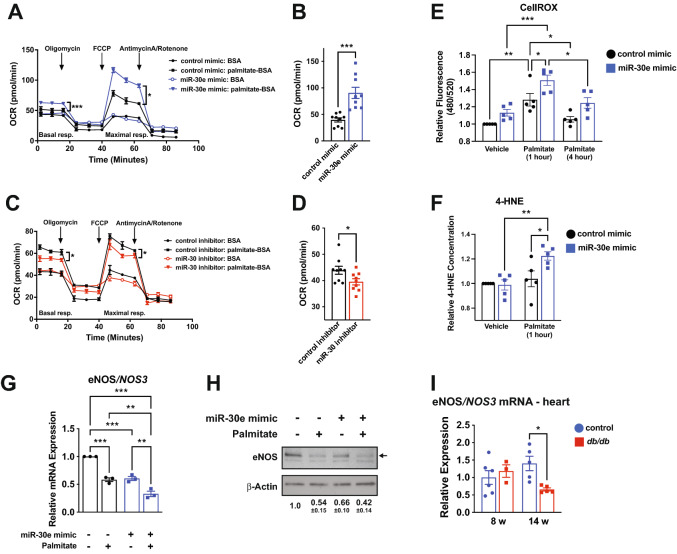
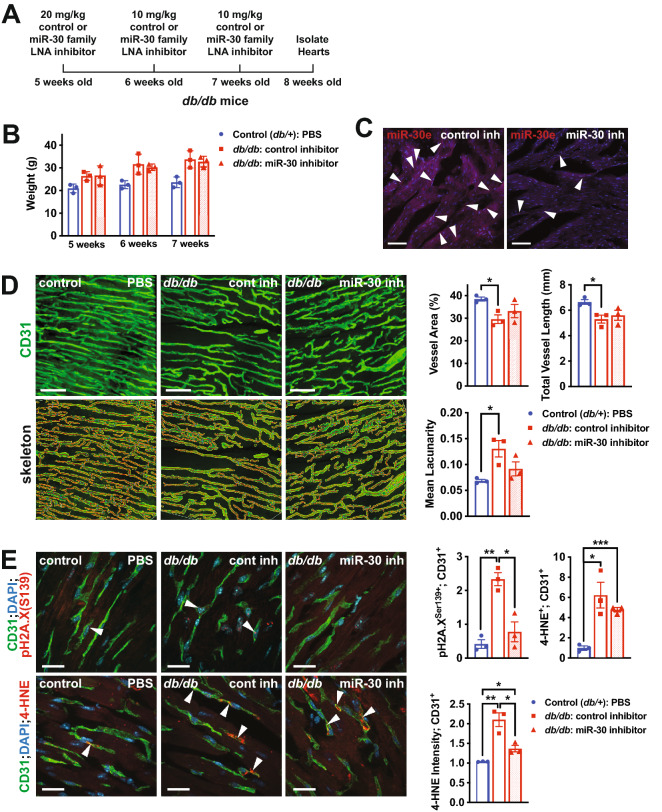
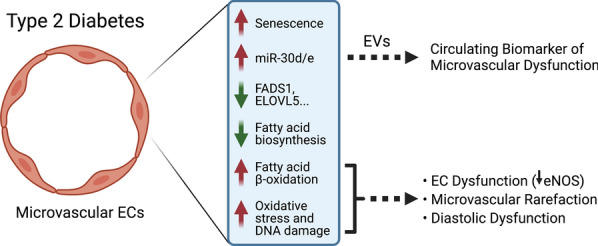
Results: We found an increase in miR-30d-5p and miR-30e-5p in circulating EVs that coincided with indices of coronary microvascular EC dysfunction (i.e., markers of oxidative stress, DNA damage/senescence) and rarefaction, and preceded echocardiographic evidence of diastolic dysfunction. These miRNAs may serve as biomarkers of coronary microvascular dysfunction as they are upregulated in ECs of the left ventricle of the heart, but not other organs, in db/db mice. Furthermore, the miR-30 family is secreted in EVs from senescent ECs in culture, and ECs with senescent-like characteristics are present in the db/db heart. Assessment of miR-30 target pathways revealed a network of genes involved in fatty acid biosynthesis and metabolism. Over-expression of miR-30e in cultured ECs increased fatty acid β-oxidation and the production of reactive oxygen species and lipid peroxidation, while inhibiting the miR-30 family decreased fatty acid β-oxidation. Additionally, miR-30e over-expression synergized with fatty acid exposure to down-regulate the expression of eNOS, a key regulator of microvascular and cardiomyocyte function. Finally, knock-down of the miR-30 family in db/db mice decreased markers of oxidative stress and DNA damage/senescence in the microvascular endothelium.
Conclusions: MiR-30d/e represent early biomarkers and potential therapeutic targets that are indicative of the development of diastolic dysfunction and may reflect altered EC fatty acid metabolism and microvascular dysfunction in the diabetic heart.
Keywords: Biomarker; Diabetes; Diastolic dysfunction; Endothelial cell; Extracellular vesicle; Heart failure with preserved ejection fraction; Microvasculature; microRNA.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Kannel WB, Hjortland M, Castelli WP. Role of diabetes in congestive heart failure: the Framingham study. Am J Cardiol. 1974;34(1):29–34. - PubMed
-
- Kannel WB, McGee DL. Diabetes and cardiovascular disease: the Framingham study. JAMA. 1979;241(19):2035–2038. - PubMed
-
- Greenberg BH, Abraham WT, Albert NM, Chiswell K, Clare R, Stough WG, et al. Influence of diabetes on characteristics and outcomes in patients hospitalized with heart failure: a report from the Organized Program to Initiate Lifesaving Treatment in Hospitalized Patients with Heart Failure (OPTIMIZE-HF) Am Heart J. 2007;154(2):277.e1–8. - PubMed
-
- Bouthoorn S, Valstar GB, Gohar A, den Ruijter HM, Reitsma HB, Hoes AW, et al. The prevalence of left ventricular diastolic dysfunction and heart failure with preserved ejection fraction in men and women with type 2 diabetes: a systematic review and meta-analysis. Diabetes Vasc Dis Res. 2018;15(6):477–493. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
