

Sex-specific differences in KCC2 localisation and inhibitory synaptic transmission in the rat hippocampus
- PMID: 35210456
- PMCID: PMC8873453
- DOI: 10.1038/s41598-022-06769-5
Sex-specific differences in KCC2 localisation and inhibitory synaptic transmission in the rat hippocampus
Abstract
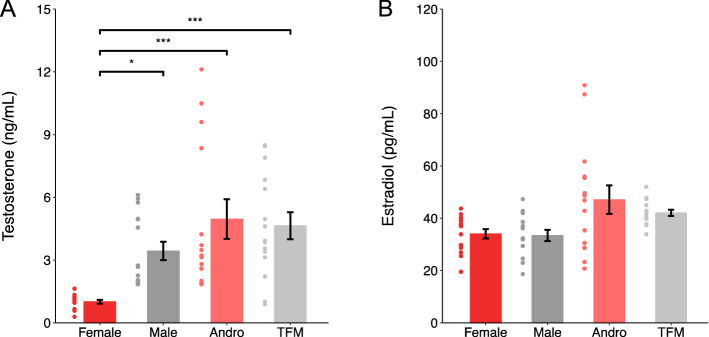
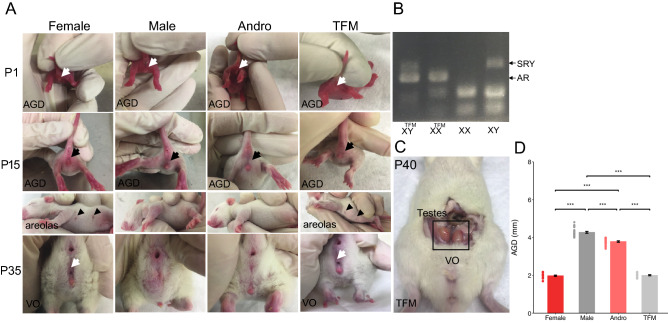
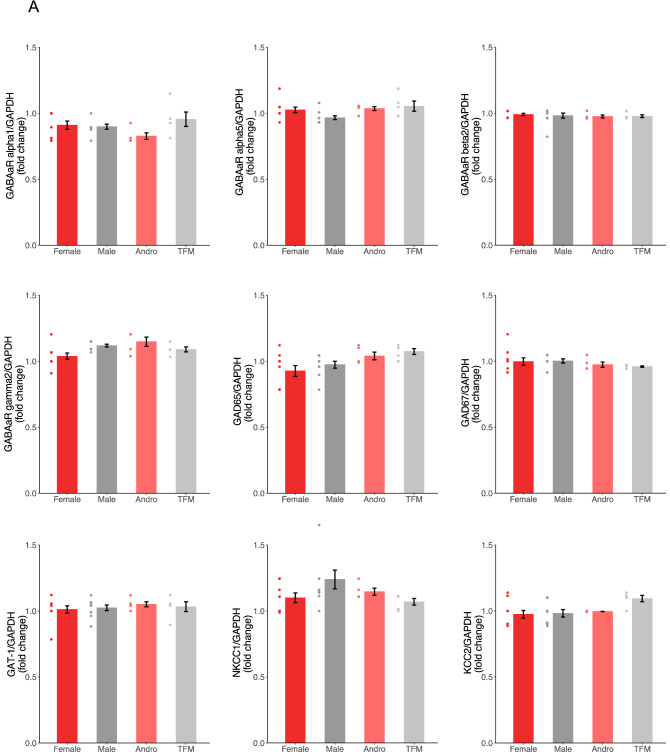
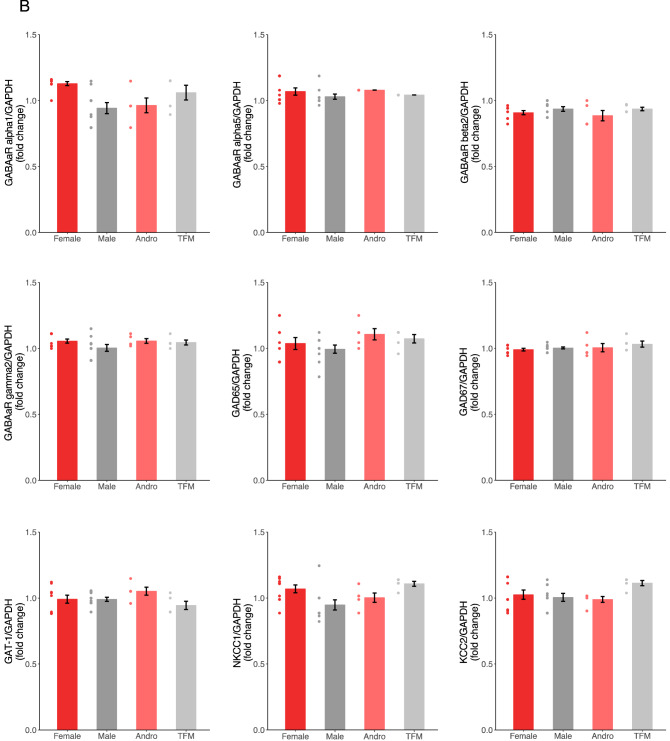
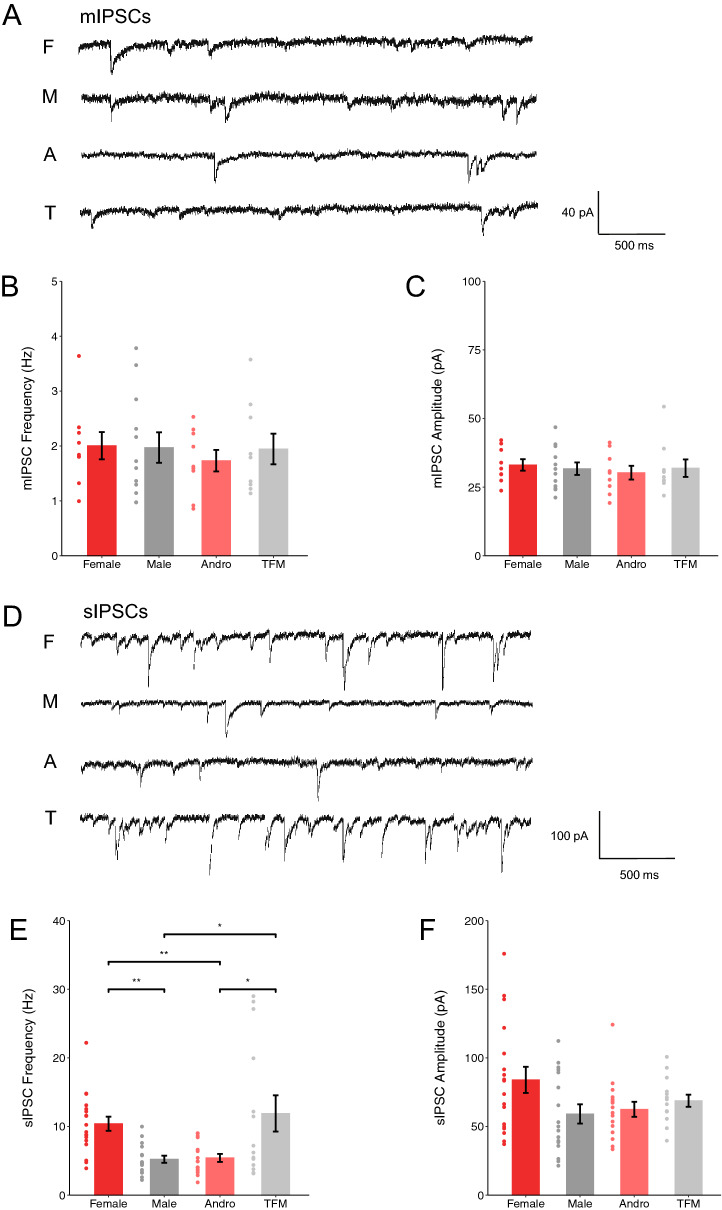
Sexual differentiation of the brain is influenced by testosterone and its metabolites during the perinatal period, when many aspects of brain development, including the maturation of GABAergic transmission, occur. Whether and how testosterone signaling during the perinatal period affects GABAergic transmission is unclear. Here, we analyzed GABAergic circuit functional markers in male, female, testosterone-treated female, and testosterone-insensitive male rats after the first postnatal week and in young adults. In the hippocampus, mRNA levels of proteins associated with GABA signaling were not significantly affected at postnatal day (P) 7 or P40. Conversely, membrane protein levels of KCC2, which are critical for determining inhibition strength, were significantly higher in females compared to males and testosterone-treated females at P7. Further, female and testosterone-insensitive male rats at P7 showed higher levels of the neurotrophin BDNF, which is a powerful regulator of neuronal function, including GABAergic transmission. Finally, spontaneous GABAergic currents in hippocampal CA1 pyramidal cells were more frequent in females and testosterone-insensitive males at P40. Overall, these results show that perinatal testosterone levels modulate GABAergic circuit function, suggesting a critical role of perinatal sex hormones in regulating network excitability in the adult hippocampus.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
