

Endothelial glycocalyx is damaged in diabetic cardiomyopathy: angiopoietin 1 restores glycocalyx and improves diastolic function in mice
- PMID: 35211778
- PMCID: PMC8960650
- DOI: 10.1007/s00125-022-05650-4
Endothelial glycocalyx is damaged in diabetic cardiomyopathy: angiopoietin 1 restores glycocalyx and improves diastolic function in mice
Abstract
Aims/hypothesis: Diabetic cardiomyopathy (DCM) is a serious and under-recognised complication of diabetes. The first sign is diastolic dysfunction, which progresses to heart failure. The pathophysiology of DCM is incompletely understood but microcirculatory changes are important. Endothelial glycocalyx (eGlx) plays multiple vital roles in the microcirculation, including in the regulation of vascular permeability, and is compromised in diabetes but has not previously been studied in the coronary microcirculation in diabetes. We hypothesised that eGlx damage in the coronary microcirculation contributes to increased microvascular permeability and hence to cardiac dysfunction.
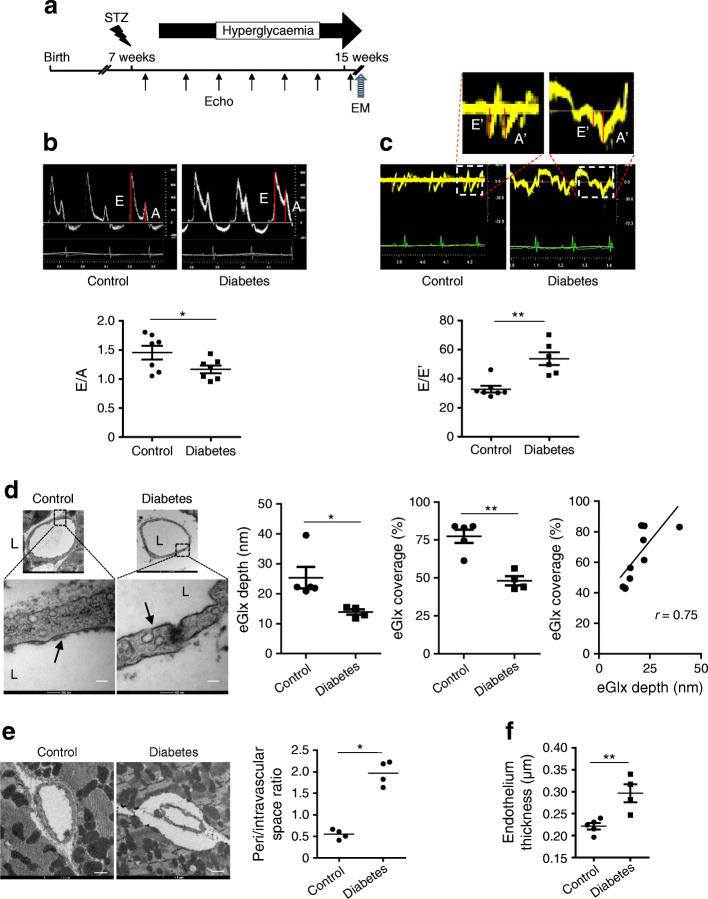
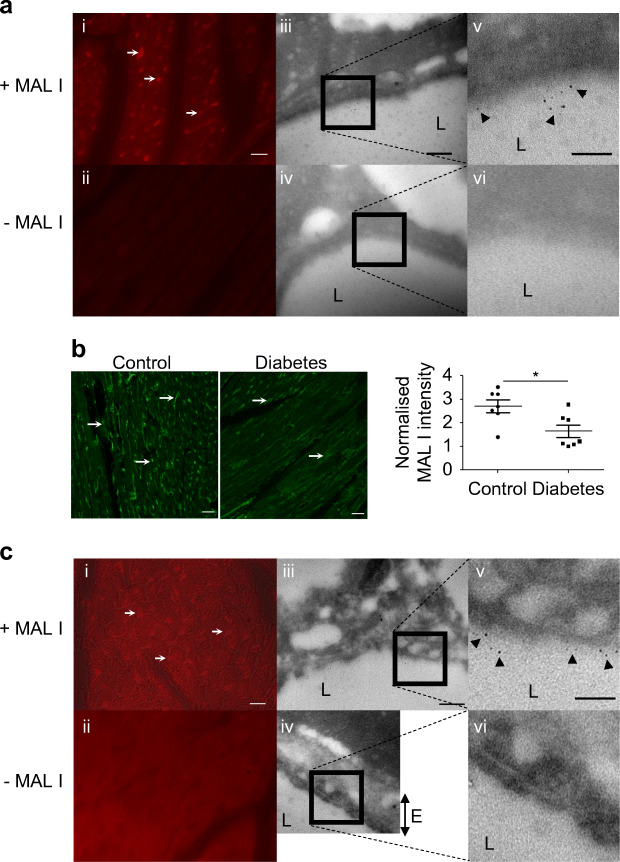
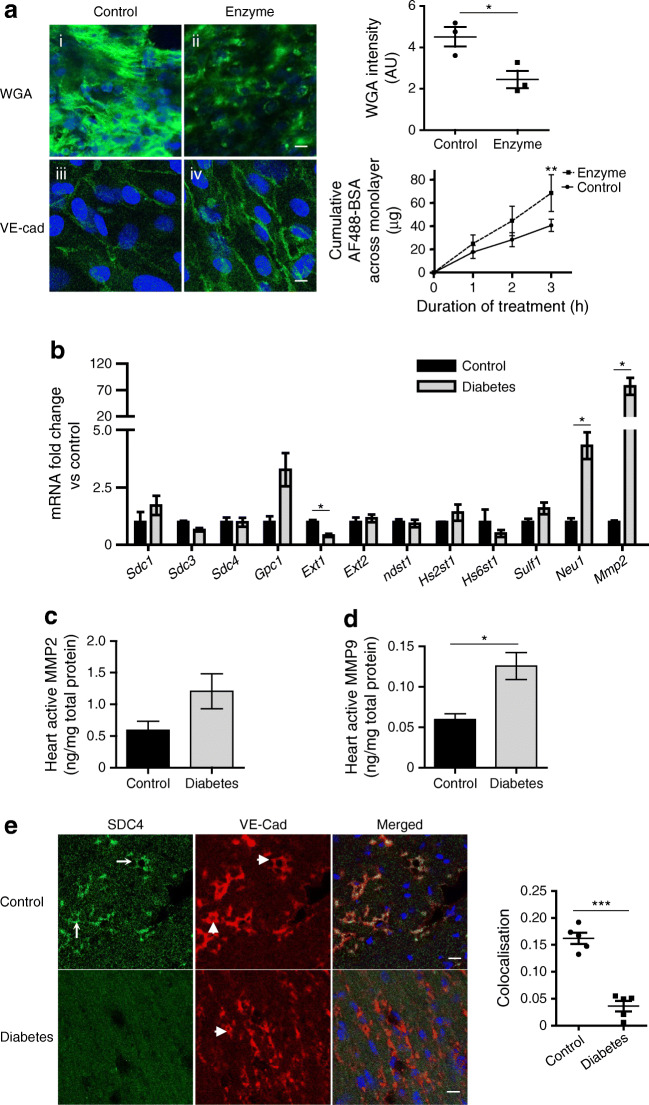
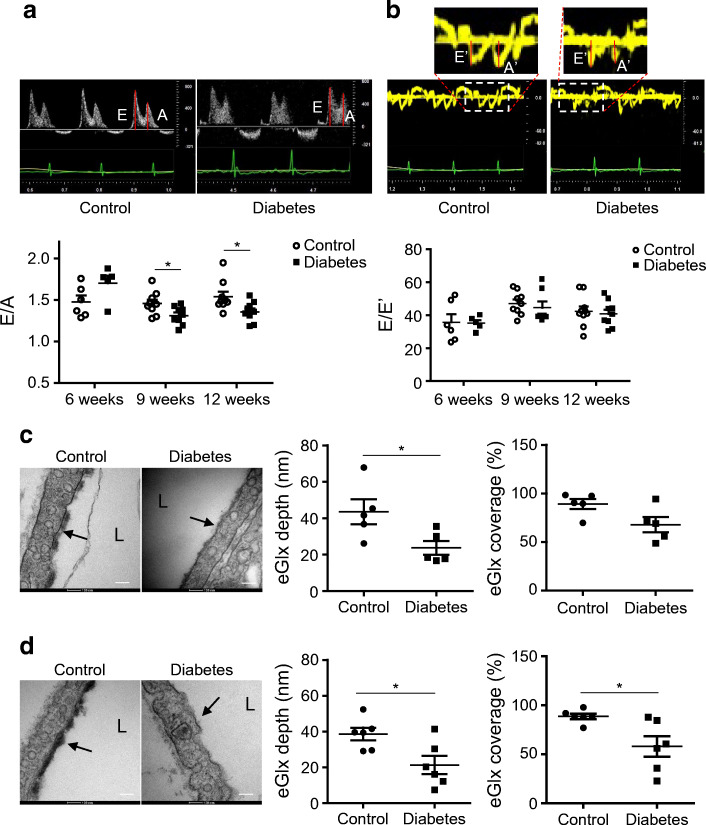
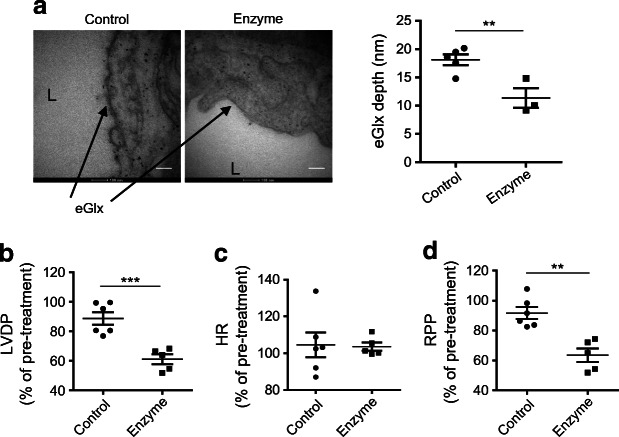
Methods: We investigated eGlx damage and cardiomyopathy in mouse models of type 1 (streptozotocin-induced) and type 2 (db/db) diabetes. Cardiac dysfunction was determined by echocardiography. We obtained eGlx depth and coverage by transmission electron microscopy (TEM) on mouse hearts perfusion-fixed with glutaraldehyde and Alcian Blue. Perivascular oedema was assessed from TEM images by measuring the perivascular space area. Lectin-based fluorescence was developed to study eGlx in paraformaldehyde-fixed mouse and human tissues. The eGlx of human conditionally immortalised coronary microvascular endothelial cells (CMVECs) in culture was removed with eGlx-degrading enzymes before measurement of protein passage across the cell monolayer. The mechanism of eGlx damage in the diabetic heart was investigated by quantitative reverse transcription-PCR array and matrix metalloproteinase (MMP) activity assay. To directly demonstrate that eGlx damage disturbs cardiac function, isolated rat hearts were treated with enzymes in a Langendorff preparation. Angiopoietin 1 (Ang1) is known to restore eGlx and so was used to investigate whether eGlx restoration reverses diastolic dysfunction in mice with type 1 diabetes.
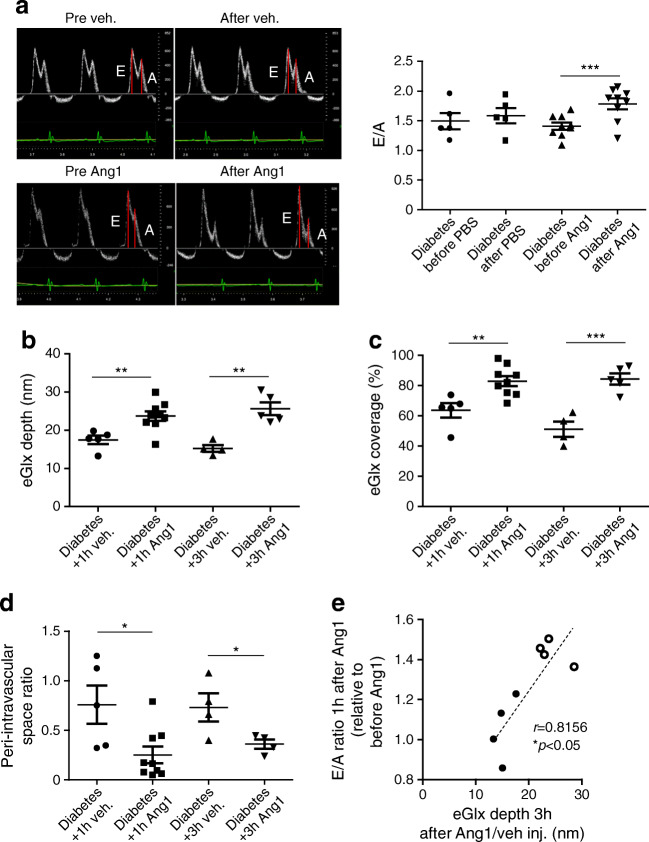
Results: In a mouse model of type 1 diabetes, diastolic dysfunction (confirmed by echocardiography) was associated with loss of eGlx from CMVECs and the development of perivascular oedema, suggesting increased microvascular permeability. We confirmed in vitro that eGlx removal increases CMVEC monolayer permeability. We identified increased MMP activity as a potential mechanism of eGlx damage and we observed loss of syndecan 4 consistent with MMP activity. In a mouse model of type 2 diabetes we found a similar loss of eGlx preceding the development of diastolic dysfunction. We used isolated rat hearts to demonstrate that eGlx damage (induced by enzymes) is sufficient to disturb cardiac function. Ang1 restored eGlx and this was associated with reduced perivascular oedema and amelioration of the diastolic dysfunction seen in mice with type 1 diabetes.
Conclusions/interpretation: The association of CMVEC glycocalyx damage with diastolic dysfunction in two diabetes models suggests that it may play a pathophysiological role and the enzyme studies confirm that eGlx damage is sufficient to impair cardiac function. Ang1 rapidly restores the CMVEC glycocalyx and improves diastolic function. Our work identifies CMVEC glycocalyx damage as a potential contributor to the development of DCM and therefore as a therapeutic target.
Keywords: Angiopoietin 1; Coronary microcirculation; Diabetes; Glycocalyx; Permeability.
© 2022. The Author(s).
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
