

Distinct roles for motor cortical and thalamic inputs to striatum during motor skill learning and execution
- PMID: 35213216
- PMCID: PMC8880788
- DOI: 10.1126/sciadv.abk0231
Distinct roles for motor cortical and thalamic inputs to striatum during motor skill learning and execution
Abstract
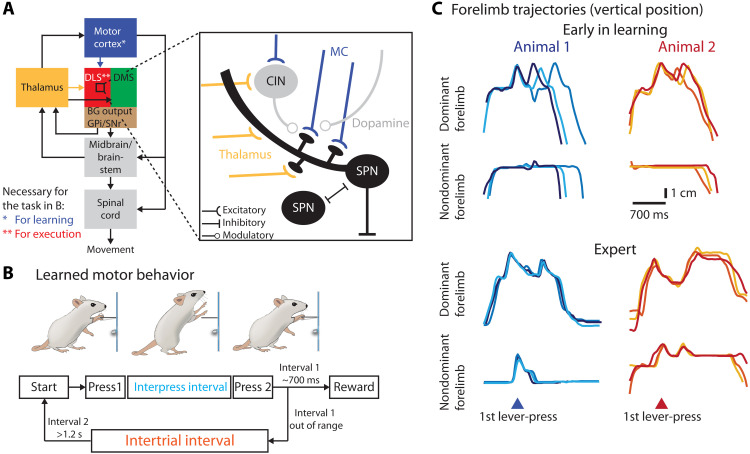
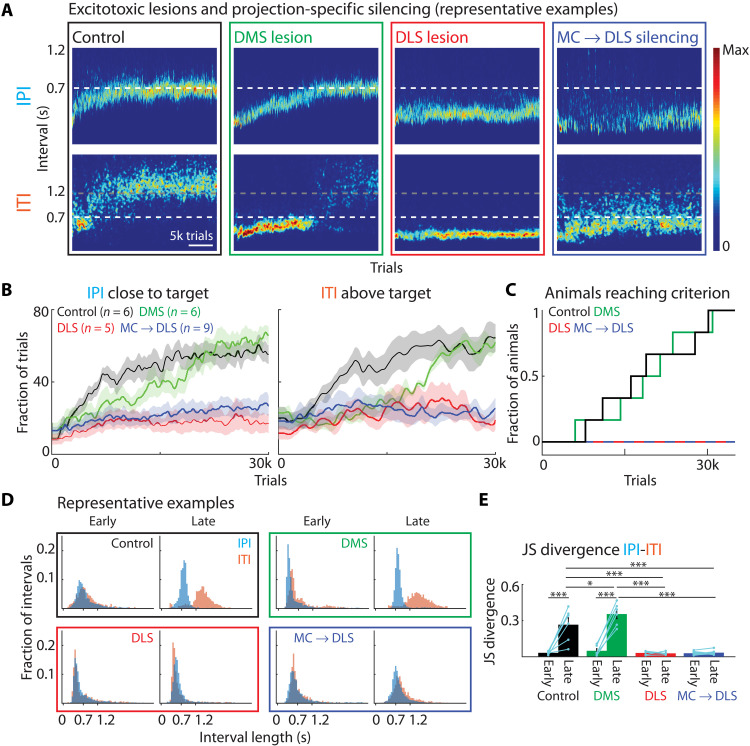
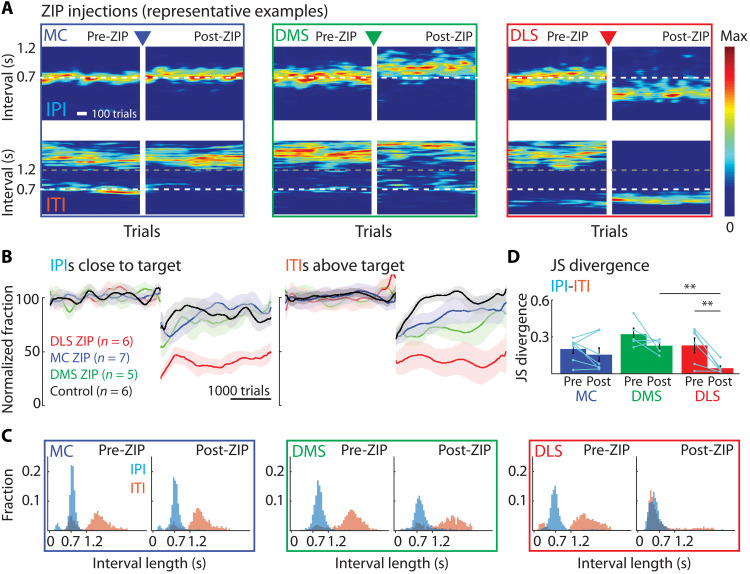
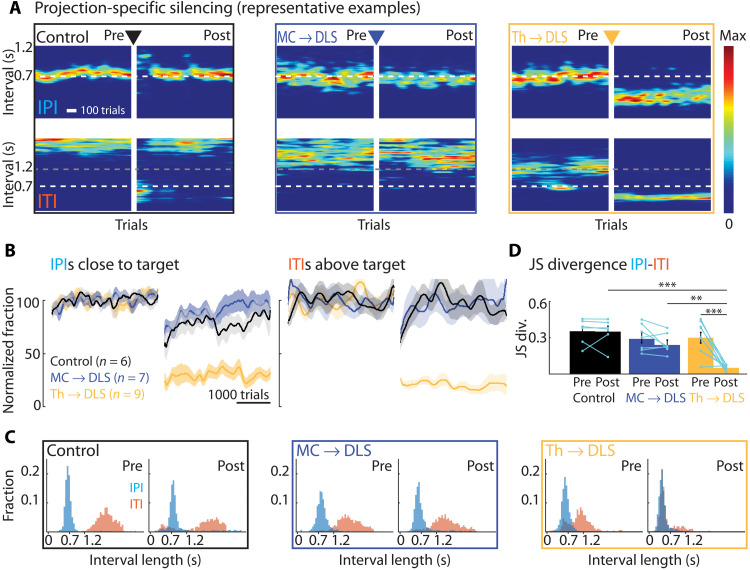
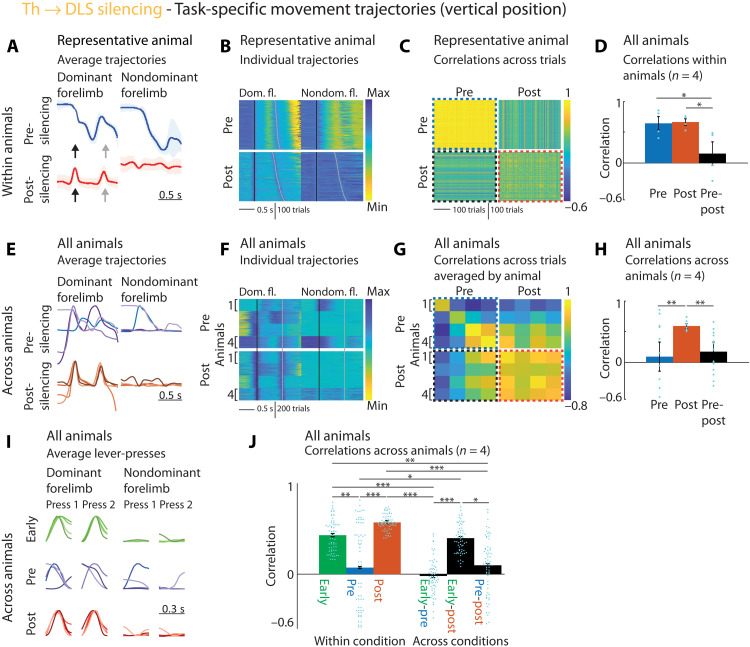
The acquisition and execution of motor skills are mediated by a distributed motor network, spanning cortical and subcortical brain areas. The sensorimotor striatum is an important cog in this network, yet the roles of its two main inputs, from motor cortex and thalamus, remain largely unknown. To address this, we silenced the inputs in rats trained on a task that results in highly stereotyped and idiosyncratic movement patterns. While striatal-projecting motor cortex neurons were critical for learning these skills, silencing this pathway after learning had no effect on performance. In contrast, silencing striatal-projecting thalamus neurons disrupted the execution of the learned skills, causing rats to revert to species-typical pressing behaviors and preventing them from relearning the task. These results show distinct roles for motor cortex and thalamus in the learning and execution of motor skills and suggest that their interaction in the striatum underlies experience-dependent changes in subcortical motor circuits.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
